RNA Interference (RNAi): Mechanism, Applications, and Therapeutic Development
What Is RNA Interference?
RNA interference (RNAi) is a conserved biological mechanism in which small double-stranded RNA molecules silence gene expression in a sequence-specific manner. The process works at the post-transcriptional level: short RNA duplexes, typically 21 to 23 nucleotides long, guide an enzymatic complex to complementary messenger RNA (mRNA) transcripts, triggering their degradation or translational repression. Because any gene can be targeted simply by designing a complementary RNA sequence, RNAi has become one of the most widely used tools in molecular biology and a rapidly expanding platform for drug development.
The phenomenon was first described in 1998 by Andrew Fire and Craig Mello, who demonstrated that injecting double-stranded RNA (dsRNA) into the nematode Caenorhabditis elegans produced potent, gene-specific silencing far more effectively than either sense or antisense RNA alone. Their observation that only a few molecules of dsRNA per cell could trigger a robust silencing response suggested an underlying catalytic mechanism. Fire and Mello received the 2006 Nobel Prize in Physiology or Medicine for this discovery. Related silencing phenomena—co-suppression in plants (observed in 1990) and quelling in the fungus Neurospora crassa—were subsequently recognized as manifestations of the same conserved pathway, indicating that RNAi arose early in eukaryotic evolution as a defense against viral infection and transposon activity.
How Does RNAi Work?
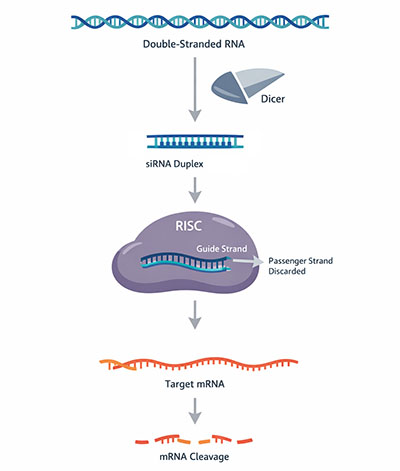
The RNAi pathway begins when long double-stranded RNA enters the cytoplasm of a cell. The RNase III family endonuclease Dicer recognizes and processes this dsRNA into short duplexes known as small interfering RNAs (siRNAs), each approximately 21 to 23 nucleotides in length with characteristic two-nucleotide 3′ overhangs. These structural features are critical for recognition by downstream silencing machinery. Dicer works with double-stranded RNA-binding proteins such as TRBP in mammals, which assist in substrate positioning and handoff of the siRNA product.
The siRNA duplex is then loaded into a multi-protein assembly called the RNA-induced silencing complex (RISC). During loading, the duplex is unwound and one strand—the passenger strand—is discarded, while the other—the guide strand—remains bound to the Argonaute protein at the catalytic core of RISC. Strand selection is governed by the thermodynamic properties of the duplex: the strand whose 5′ end is less stably base-paired is preferentially retained as the guide. In humans, Argonaute 2 (Ago2) is the only family member with endonucleolytic (slicer) activity, capable of cleaving target mRNA between positions 10 and 11 relative to the guide strand's 5′ end.
Once loaded, the guide-strand–RISC complex scans cytoplasmic mRNAs for sequences complementary to the guide. When near-perfect complementarity is found, Ago2 cleaves the target mRNA, and the resulting fragments are rapidly degraded by cellular exonucleases. Because RISC is not consumed in this reaction, a single activated complex can sequentially destroy multiple copies of the target transcript, explaining the catalytic potency that Fire and Mello originally observed. A related but mechanistically distinct pathway is employed by endogenous microRNAs (miRNAs), which bind with only partial complementarity and typically repress translation rather than triggering mRNA cleavage.
Biological Functions of RNAi
RNAi is not merely a laboratory tool—it serves essential regulatory and defensive functions across eukaryotic biology. One of its most ancient roles is antiviral defense. Many viruses produce double-stranded RNA intermediates during replication, and the RNAi pathway recognizes and destroys these molecules, limiting viral propagation. This antiviral function is particularly prominent in plants and invertebrates, where RNAi acts as a primary innate immune mechanism against RNA viruses. In mammals, the interferon response largely supplants RNAi as the dominant antiviral defense, although RNAi components remain active and may contribute to the control of certain viral infections.
RNAi also plays a central role in genome defense by suppressing the activity of transposable elements—parasitic DNA sequences that can replicate and insert themselves at new positions in the genome, potentially disrupting gene function. In the germline of animals, a specialized branch of the RNAi machinery involving PIWI-interacting RNAs (piRNAs) silences transposons and is essential for maintaining genome integrity across generations. In plants and fungi, RNAi-directed DNA methylation provides an additional layer of transposon control at the transcriptional level, linking the pathway to epigenetic regulation.
Beyond defense, the endogenous miRNA arm of the RNAi pathway is a master regulator of gene regulation during development and tissue homeostasis. The human genome encodes more than 2,500 mature miRNAs, which collectively regulate the expression of over 60% of all protein-coding genes by fine-tuning cell differentiation, proliferation, apoptosis, and metabolic balance.
RNAi as a Research Tool
The demonstration in 2001 by Elbashir and colleagues that synthetic 21-nucleotide siRNA duplexes could silence specific genes in mammalian cells without triggering the nonspecific interferon response associated with longer dsRNAs transformed RNAi from a biological curiosity into a practical laboratory tool. Researchers can now design siRNAs against virtually any gene of interest, introduce them into cultured cells by transfection, and observe the phenotypic consequences of gene knockdown within 24 to 72 hours. This approach is far faster and less expensive than generating knockout animals or stable mutant cell lines.
Genome-wide RNAi screens, in which libraries of siRNAs targeting every gene in the genome are tested systematically, have been used to identify genes involved in cell division, viral entry, drug resistance, and signal transduction. In C. elegans and Drosophila melanogaster, RNAi can be delivered simply by feeding organisms bacteria that express the corresponding dsRNA, enabling large-scale genetic screens at minimal cost. Short hairpin RNAs (shRNAs), expressed from DNA vectors and processed into siRNAs by Dicer, provide an alternative that allows stable, long-term gene silencing in cell lines and transgenic animals.
Therapeutic Applications of RNAi
Translating RNAi from laboratory tool to clinical therapy required solving two fundamental challenges: delivering siRNA molecules to the correct tissue and protecting them from rapid degradation by serum nucleases. Naked siRNA is cleared from the bloodstream within minutes, and its large size (approximately 14 kDa) and negative charge prevent passive diffusion across cell membranes. Two delivery strategies have reached commercial success. The first, used in patisiran (Onpattro), encapsulates siRNA in lipid nanoparticles (LNPs) that protect the cargo and facilitate uptake by hepatocytes through apolipoprotein E-mediated endocytosis. The second, used in six subsequent approvals, conjugates chemically stabilized siRNA to N-acetylgalactosamine (GalNAc), a sugar ligand that binds with high affinity to the asialoglycoprotein receptor on the surface of liver cells.
Patisiran, approved by the FDA in August 2018, was the first RNAi therapeutic to reach the market. It targets transthyretin (TTR) mRNA in the liver, reducing the production of misfolded TTR protein that accumulates as amyloid deposits in the peripheral nerves and heart of patients with hereditary transthyretin-mediated amyloidosis (hATTR). Six additional siRNA drugs have since received FDA approval: givosiran (2019) for acute hepatic porphyria, lumasiran (2020) for primary hyperoxaluria type 1, inclisiran (2021) for lowering LDL cholesterol, vutrisiran (2022) for hATTR polyneuropathy, nedosiran (2023) for primary hyperoxaluria, and fitusiran (2025) for prophylaxis of bleeding episodes in hemophilia A or B. Also in 2025, vutrisiran's indication was expanded to include ATTR amyloidosis with cardiomyopathy, marking the first RNAi therapeutic approved to reduce cardiovascular death.
All seven approved siRNA drugs target hepatocytes, reflecting the relative maturity of liver-directed delivery systems. Extending RNAi therapeutics to other tissues—the central nervous system, lungs, kidneys, and solid tumors—remains a major research focus. Approaches under investigation include antibody–siRNA conjugates for tissue-specific targeting, exosome-mediated delivery, and polymer-based nanoparticles engineered for pulmonary or intrathecal administration.
Comparison of RNAi-Based Approaches
Several molecular tools exploit the RNAi pathway, each suited to particular research or therapeutic contexts. The table below compares the most commonly used RNAi trigger formats.
| Feature | Synthetic siRNA | Short Hairpin RNA (shRNA) | miRNA Mimic |
|---|---|---|---|
| Structure | 21–23 nt duplex with 3′ overhangs | Stem-loop expressed from a DNA vector | Synthetic duplex mimicking endogenous miRNA |
| Delivery | Transfection, LNP, or GalNAc conjugate | Viral vector or plasmid | Transfection or LNP |
| Duration of silencing | Transient (3–7 days in dividing cells) | Stable (weeks to permanent with integration) | Transient (3–7 days) |
| Target specificity | Single gene (perfect complementarity) | Single gene (processed to siRNA by Dicer) | Multiple genes (partial complementarity) |
| Primary mechanism | mRNA cleavage by Ago2 | mRNA cleavage by Ago2 | Translational repression and mRNA destabilization |
| Clinical use | Seven FDA-approved drugs | Research tool; limited clinical use | Early-stage clinical trials |
The choice among these formats depends on the application. Synthetic siRNAs dominate therapeutic development because of their defined chemical composition and amenability to stabilizing modifications. shRNAs are preferred when sustained knockdown is needed in cell-based studies or animal models, while miRNA mimics and inhibitors are used to modulate endogenous miRNA pathways.
Limitations and Challenges
Off-target effects remain a persistent concern. The guide strand can bind to unintended mRNAs sharing partial sequence complementarity, particularly in the seed region (positions 2 to 8 from the 5′ end), mimicking miRNA-like activity and altering expression of unrelated genes. Careful siRNA design using bioinformatic algorithms, combined with chemical modifications such as 2′-O-methylation of position 2, can substantially reduce seed-mediated off-target silencing but cannot eliminate it entirely.
Delivery to extrahepatic tissues remains the most significant barrier to expanding the clinical reach of RNAi therapeutics. The liver benefits from a unique combination of fenestrated endothelium, high blood flow, and receptor systems (particularly the asialoglycoprotein receptor) that facilitate efficient siRNA uptake. Other organs lack comparable receptor-mediated entry pathways, and biological barriers such as the blood–brain barrier, the pulmonary mucus layer, and the dense extracellular matrix of solid tumors present additional obstacles. Even with current delivery systems, endosomal escape remains inefficient, with estimates suggesting that fewer than 1 to 2% of endocytosed siRNA molecules reach the cytoplasm where RISC operates.
Immune stimulation by synthetic RNA duplexes can also complicate therapeutic development. Double-stranded RNA is recognized by innate immune sensors including Toll-like receptors 3, 7, and 8 and the cytoplasmic sensor RIG-I, potentially triggering interferon production and inflammatory cytokine release. Strategic incorporation of 2′-O-methyl and 2′-fluoro modifications throughout the duplex, a strategy employed in all clinically approved siRNAs, effectively suppresses these responses.
Future Perspectives
The trajectory of RNAi therapeutics is shaped by two converging trends: advances in delivery technology and the maturation of GalNAc-conjugate chemistry. The commercial success of twice-yearly dosing with inclisiran for cholesterol management has demonstrated that RNAi can compete with established drug classes in large patient populations, not only in rare diseases. As delivery platforms capable of reaching the central nervous system, kidneys, and lungs advance through clinical development, the addressable patient population for RNAi-based medicines is expected to grow substantially. Antibody–siRNA conjugates that exploit tissue-specific surface receptors for targeted delivery are among the most promising extrahepatic approaches, with several candidates in early clinical trials for muscle diseases and solid tumors.
Artificial intelligence and computational design are accelerating siRNA lead discovery. Machine learning models trained on large datasets of siRNA activity and off-target profiles can now predict potent, specific guide sequences more reliably than rule-based algorithms. Integration of RNAi with complementary modalities—antisense oligonucleotides, mRNA therapeutics, and CRISPR-based gene editing—is creating combination strategies in which transient RNAi-mediated silencing works alongside permanent genetic correction. In personalized medicine, the rapid design cycle of siRNA—from target sequence to candidate molecule in days—positions RNAi as a platform uniquely suited to individualized therapy.
Frequently Asked Questions
What is the difference between siRNA and miRNA? Small interfering RNA (siRNA) and microRNA (miRNA) both operate through the RNAi pathway and associate with the RISC complex, but they differ in origin and targeting behavior. siRNAs are derived from long double-stranded RNA precursors and typically bind with perfect complementarity to a single target mRNA, triggering its cleavage and degradation. miRNAs are encoded by endogenous genes, processed from hairpin-shaped precursors, and usually bind with partial complementarity to the 3′ untranslated region of target mRNAs, resulting in translational repression rather than cleavage. A single miRNA can regulate hundreds of different transcripts, whereas siRNAs are generally specific to one target.
Why do all FDA-approved siRNA drugs target the liver? The liver's anatomy and physiology make it the most accessible organ for siRNA delivery. Hepatocytes have fenestrated endothelium that allows nanoparticles to pass from the bloodstream into liver tissue, and they express high levels of the asialoglycoprotein receptor, which binds N-acetylgalactosamine (GalNAc) conjugates with high affinity. GalNAc-conjugated siRNAs exploit this receptor for efficient and specific uptake into hepatocytes. Delivering siRNAs to other organs such as the brain, lungs, or kidneys remains a major challenge because these tissues lack comparably efficient receptor-mediated uptake pathways and present additional biological barriers.
Can RNA interference permanently alter the genome? No. RNA interference acts at the post-transcriptional level by degrading messenger RNA or blocking its translation into protein. It does not modify DNA sequence or structure. The silencing effect is transient because the target gene remains intact and continues to be transcribed. Once the siRNA molecules are cleared or diluted through cell division, gene expression returns to normal levels. This reversibility distinguishes RNAi from gene-editing technologies such as CRISPR-Cas9, which make permanent changes to the DNA.
How often do patients need to take RNAi drugs? Dosing frequency varies by drug and delivery platform. Patisiran, the first approved siRNA therapeutic, is administered as an intravenous infusion every three weeks. Newer GalNAc-conjugated siRNAs require far less frequent dosing: inclisiran is given by subcutaneous injection twice yearly after an initial dose and a three-month follow-up, and vutrisiran is administered subcutaneously once every three months. The extended dosing intervals of GalNAc-conjugated siRNAs reflect their efficient hepatocyte uptake, metabolic stability conferred by chemical modifications, and sustained pharmacodynamic activity.
Is RNAi being used in agriculture? Yes. RNAi-based approaches have been applied in agriculture both through transgenic crops and through topical application of double-stranded RNA. Transgenic crops engineered to express dsRNA targeting essential pest genes have been developed, including corn varieties that produce dsRNA against the western corn rootworm. Spray-on dsRNA products are also under investigation as alternatives to conventional chemical pesticides, offering species-specific pest control with potentially reduced environmental impact. RNAi has additionally been used to modify crop traits such as reduced browning in apples and improved nutritional profiles.
Further Reading
Annual Review of Biophysics, Molecular Mechanisms of RNA Interference
Nature Reviews Drug Discovery, The Current State and Future Directions of RNAi-Based Therapeutics
Signal Transduction and Targeted Therapy, Therapeutic siRNA: State of the Art
