What is DNA Origami? Structure, Design and Applications
What is DNA Origami?
In one sentence: DNA origami is a self-assembly technique that folds a long single-stranded DNA molecule into precisely defined nanoscale shapes using hundreds of short, computer-designed staple strands.
DNA origami exploits sequence-specific Watson–Crick base pairing to fold a long single-stranded DNA molecule, called the scaffold, into precisely defined two- or three-dimensional shapes. Hundreds of short synthetic oligonucleotides, called staple strands, are designed so that each binds two or more separate regions of the scaffold and pulls them together. When scaffold and staples are mixed in a magnesium-containing buffer and slowly cooled from around 65–85 °C to room temperature, the entire structure self-assembles in a single annealing step into the target shape, with nanometre-scale positional control that is often on the order of a few nanometres, depending on design, flexibility and measurement method. The technique is a representative example of bottom-up nanotechnology at the nanoscale.
The method was introduced by Paul Rothemund at the California Institute of Technology in a 2006 Nature paper (Folding DNA to create nanoscale shapes and patterns) that folded a 7-kilobase viral genome into shapes such as squares, disks, triangles, smiley faces and a map of the western hemisphere, all roughly 100 nm across with a spatial resolution of about 6 nm. The approach built on three decades of structural DNA nanotechnology dating to Nadrian Seeman's immobile Holliday junctions in 1982, but it simplified construction dramatically by replacing many carefully tuned multi-strand tile designs with a scaffold-plus-staples strategy. Modern designs routinely use the M13mp18 bacteriophage genome (7,249 nucleotides) with about 200 staple strands of 20 to 60 nucleotides each, and yields of correctly folded structures can reach 60–90% for many simple or well-optimised designs under standard conditions.
DNA origami complements other nucleic-acid self-assembly methods such as DNA tiles, DNA bricks, single-stranded tiles and RNA origami. Its central appeal is addressability: every base position on the folded object is known, so chemical groups, dyes, proteins and other nanoparticles can be attached at user-specified locations with nanometre-scale precision, typically on the order of a few nanometres. This has made the technique a programmable platform for molecular nanotechnology, with applications ranging from drug delivery and biosensing to nanoscale photonics, single-molecule biophysics and the prototyping of molecular machines.
Key takeaways:
- DNA origami folds a long single-stranded DNA scaffold into a designed shape using hundreds of short staple strands that bind specific regions of the scaffold together.
- The technique was introduced in 2006 and routinely produces 2D and 3D structures roughly 20–150 nm in size, with sub-10 nm spatial addressability across the entire object.
- Computer-aided design tools such as caDNAno and scadnano automate the conversion of a target shape into a list of staple sequences.
- Applications include drug and gene delivery, vaccine scaffolds, biosensors, DNA-PAINT super-resolution imaging, plasmonic nanophotonics, single-molecule biophysics and molecular machines.
| Quick fact | Typical value or feature |
|---|---|
| Building blocks | Long scaffold strand plus short staple strands |
| Common scaffold | M13mp18, 7,249 nucleotides |
| Typical size | About 20–150 nm for many single-scaffold objects |
| Addressability | Nanometre-scale placement of dyes, proteins, nanoparticles and other groups |
| Assembly | Thermal annealing in a magnesium-containing buffer |
| Main strengths | Programmability, addressability and molecular-scale patterning |
| Main limits | Physiological stability, purification, cost and scale-up |
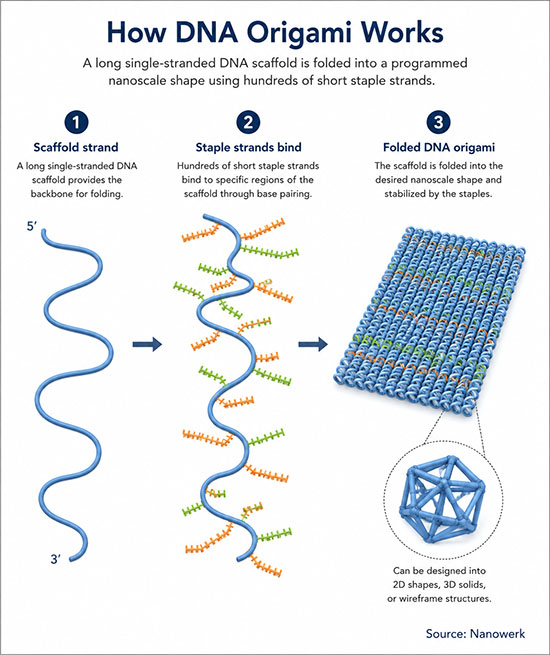
How DNA Origami Works
At the heart of DNA origami lies the geometry of the DNA double helix. Two complementary single strands wind around each other with a pitch of about 10.5 base pairs per turn, a rise of 0.34 nm per base, and a diameter of roughly 2 nm. By choosing which bases on a staple pair with which on the scaffold, a designer decides where on the long strand two regions should meet, where the helix should bend, and where it should cross to a neighbouring helix. The resulting structure is built from many parallel double-helix segments joined at well-defined crossover points, where staple strands hop between adjacent helices.
A typical 2D origami is laid out as a flat array of parallel double helices, with the scaffold zigzagging back and forth across the design and the staples acting as cross-stitches that lock neighbouring helices together. 3D origami extends the idea into a honeycomb or square lattice of helices and produces block-like, rod-like, or curved objects that are several helix diameters thick. Stiffness emerges from base stacking, hydrogen bonding and steric packing within the lattice. DNA origami can therefore be viewed as a programmable form of supramolecular assembly and molecular assembly, in which base-pairing rules act as a routing code for guiding helices through a target shape.
Designing and Folding a DNA Origami Structure
Designing an origami typically begins with a target shape sketched as a set of parallel helices on a regular lattice. Software tools — caDNAno, scadnano, vHelix, Adenita and DAEDALUS, among others — convert the geometry into a list of crossover positions and output the sequences of the staples that, together with a chosen scaffold, will fold into the target. caDNAno is the most widely used package and underlies a large fraction of the published 3D designs. For wireframe and curved designs, automated algorithms based on graph theory generate staple sets directly from a polygonal mesh with no manual editing of crossovers.
The designed staples are then synthesised commercially as short oligonucleotides, mixed with the scaffold in a magnesium-containing buffer such as TAE-Mg, and annealed. For simple 2D objects, a thermal ramp of a few hours from 65 °C to room temperature is sufficient; multilayer 3D objects can require ramps of one to several days. Folded structures are usually purified by agarose-gel electrophoresis, polyethylene-glycol precipitation, or rate-zonal centrifugation, which separate fully formed origami from misfolded species and excess staples.
Characterisation relies on atomic force microscopy for surface-deposited 2D objects, transmission electron microscopy with negative staining for 3D structures, and cryo-electron microscopy for high-resolution snapshots of native-state assemblies. Cryo-electron microscopy has resolved internal helical features of larger origami at sub-nanometre detail, while high-speed atomic force microscopy can directly visualise their conformational dynamics in solution.
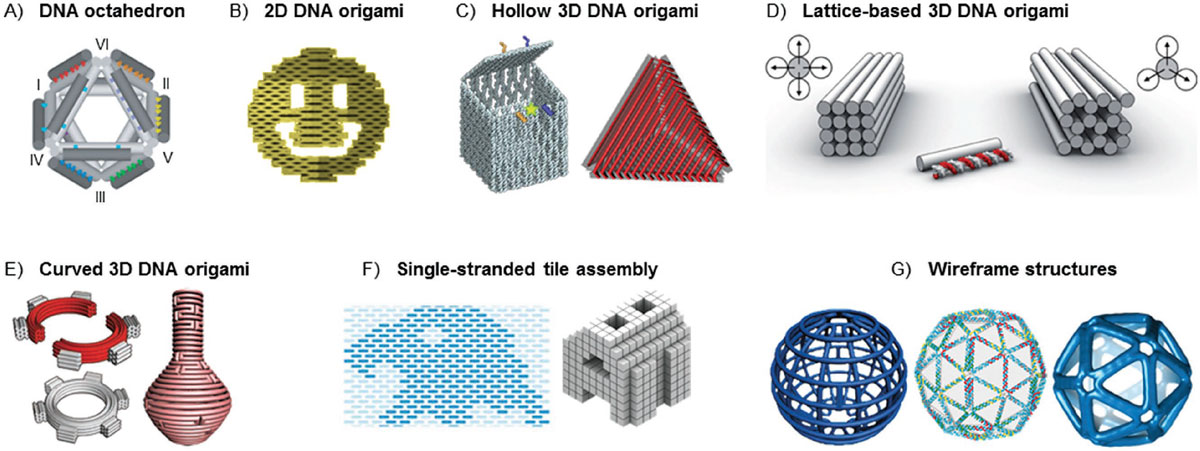
Major DNA Origami Architectures and Related Nucleic-Acid Self-Assembly Methods
Since the original flat designs in 2006, DNA origami has expanded into several distinct architectural families with different fabrication routes and target applications. The table summarises the major scaffolded DNA origami architectures and closely related nucleic-acid self-assembly methods.
| Variant | Structural principle | Typical size | Mechanical character | Typical uses |
|---|---|---|---|---|
| Classic 2D origami | Parallel double helices in a flat array; staples bridge neighbours | 70–150 nm across, ≈ 2 nm thick | Flexible mat | Surface patterning, nanoscale rulers, DNA-PAINT docking |
| Multilayer 3D origami | Helices packed on a honeycomb or square lattice | 20–100 nm in each dimension | Rigid block | Drug carriers, nanopores, mechanical biophysics |
| Wireframe / polyhedral origami | Polygonal mesh; each edge is a small helix bundle | 50–200 nm | Light, salt-tolerant cage | In vivo delivery, vaccine scaffolds, plasmonic templates |
| DNA bricks / single-stranded tiles | Short overlapping single strands; no central scaffold | Up to about 50 nm | Modular, fully addressable | Sub-100 nm shapes with very high modularity |
| RNA origami | Single co-transcriptionally folding RNA strand | 10–200 nm | Genetically encodable | Synthetic cells, in vivo expression of nanostructures |
Wireframe designs have become particularly important for biomedical work because their open mesh can remain intact under lower-magnesium and physiological-like conditions where compact multilayer origami may destabilise. Multilayer designs, on the other hand, deliver the highest mechanical stiffness and are preferred when the application requires a rigid platform with precise internal geometry, such as a membrane-spanning nanopore or a mechanical force sensor.
Functionalising DNA Origami
Because each staple position can be individually modified, DNA origami offers extraordinary control over what is attached to a structure and where. Single staples can be ordered with a 5′ or 3′ chemical modification — a fluorescent dye, a biotin, an azide, a thiol, an amine, or a click-chemistry handle — that places that group at a specific point on the folded object. Proteins, antibodies, aptamers, peptides, drugs, gold nanoparticles, quantum dots, and lipid moieties have all been positioned on DNA origami with nanometre-scale positional control, often on the order of a few nanometres depending on design, flexibility and measurement method.
This site-specific functionalisation underpins most of the applications below. When metal nanoparticles are attached at defined positions, their plasmonic near fields interact in ways that depend on spacing and orientation, allowing researchers to build chiral plasmonic structures, colour-tunable substrates and surface-enhanced Raman platforms. Pairs of fluorescent dyes positioned at known spacings serve as reference rulers for distance-dependent measurements such as Förster resonance energy transfer. Cargo molecules tethered inside a hollow origami can be released conditionally by signals that disrupt staples or open a designed lid.
Applications of DNA Origami
DNA origami is less a single product technology than a programmable platform: its value comes from positioning molecules, particles, pores and mechanical elements with nanometre-scale control.
Drug delivery, vaccines and gene therapy
A central application is the use of DNA origami as a programmable nanocarrier for drugs, nucleic acids and immunostimulatory molecules, complementing lipid-based vehicles such as liposomes and lipid nanoparticles. The intercalator doxorubicin loads efficiently into double-stranded regions and has been delivered to tumours in mice with reduced systemic toxicity. Logic-gated origami nanorobots carry payloads that release only when specific aptamer-bound triggers are present, a step toward genuinely targeted drug delivery. Wireframe origami decorated with multiple copies of antigens have been used as vaccine scaffolds that mimic the spatial arrangement of epitopes on viral surfaces and elicit strong antibody responses, an active area at the interface of nanomedicine and nanobiotechnology.
Single-molecule biophysics and biosensing
DNA origami platforms have become a widely used tool in single-molecule biophysics. By placing two molecules at precisely known separations, researchers measure how DNA-binding proteins search and bend their targets, how enzymes interact in cascades, and how membrane proteins respond to controlled forces or curvatures. Membrane-spanning DNA origami nanopores, short barrels of DNA threaded through a lipid or solid-state membrane, extend the reach of nanopore measurements to larger analytes that cannot pass through biological pores alone, complementing established nanopore DNA sequencing technology. In nanobiosensor designs, structure-switching origami change their conformation in response to a chosen target and produce fluorescence, plasmonic or electrochemical readouts.
Super-resolution imaging with DNA-PAINT
DNA-PAINT, a super-resolution microscopy method that uses transiently binding fluorescent imager strands to record blinking events at fixed origami docking sites, has become a standard tool for nanometre-accurate fluorescence imaging. Origami rulers carrying dyes at precisely known separations are sold as calibration standards for single-molecule localisation microscopy and underpin many recent advances in single-molecule imaging.
Nanophotonics and molecular machines
Origami-templated arrays of gold or silver nanoparticles produce chiral metamaterials, plasmonic colour pixels and surface-enhanced Raman substrates with periodicity set by the DNA design rather than by lithography. Reconfigurable origami structures, driven by DNA strand-displacement reactions, pH, or light, function as molecular switches, rotors, hinges and walkers — laboratory prototypes of artificial molecular motors and nanomachines that move under environmental control. These efforts overlap with broader work on programmable nanobiomaterials that respond to specific biological signals.
Challenges and Future Perspectives
Several practical limitations continue to shape the field. Classical DNA origami is sensitive to low magnesium concentrations and to nucleases, so structures placed in physiological media often need protective coatings. Oligolysine–polyethylene glycol layers, lipid bilayer wraps, virus-capsid-like protein cages and silica mineralisation have all been used to extend in vivo lifetime from minutes to hours or even days. Wireframe designs avoid some of these issues by tolerating much lower salt and by exposing fewer free staple ends, which is one reason wireframe designs are increasingly prominent in biomedical demonstrations.
Scale is another bottleneck. A standard laboratory annealing reaction produces sub-microgram amounts of folded origami. Biotechnological production of scaffold and staple strands has shown that DNA origami can be made in macroscopic quantities, moving the field beyond early microgram-scale laboratory reactions and lowering material costs. Design automation continues to expand: graph-neural-network methods reported in 2024 can rapidly predict the three-dimensional conformation of DNA origami assemblies, while newer machine-learning approaches are beginning to address stability in physiological media and reduce the empirical trial-and-error cycle.
The field is moving from static demonstrations to functional devices that operate in cellular and clinical environments. Active areas include vaccine and immunotherapy scaffolds, gene-delivery vehicles based on DNA origami-condensed plasmids, integration of DNA origami with semiconductor lithography to build hybrid molecular-electronic devices, and the use of RNA origami as a route to in vivo, genetically encoded nanostructures expressed directly inside cells.
FAQ: DNA Origami
Who invented DNA origami?
DNA origami was introduced by Paul Rothemund at Caltech in 2006. It built on earlier structural DNA nanotechnology pioneered by Nadrian Seeman, especially immobile Holliday junctions and tile-based assemblies, but simplified the design problem by folding one long scaffold with many short staple strands.
What is the difference between DNA origami and DNA nanotechnology?
DNA nanotechnology is the broader field that uses DNA as a programmable structural material. DNA origami is one method within that field, defined by folding a long scaffold strand with many short staple strands. Related approaches include DNA tiles, DNA bricks, single-stranded tiles, RNA origami and strand-displacement circuits.
How big are DNA origami structures?
Single-scaffold DNA origami structures are commonly tens to hundreds of nanometres in size. Classic 2D designs are often 70–150 nm across and about 2 nm thick, while multilayer 3D designs are commonly 20–100 nm in each dimension. Larger assemblies can be made with longer scaffolds, multiple scaffolds or hierarchical assembly.
Why does DNA origami use the M13 phage genome as a scaffold?
The M13mp18 phage genome is useful because it is a naturally single-stranded, circular DNA molecule of 7,249 nucleotides that can be amplified in E. coli at relatively low cost. Its well-characterised sequence and convenient length made it the standard scaffold for many early and current DNA origami designs.
Are DNA origami structures stable in the body?
Unprotected DNA origami can lose structure or be degraded in physiological media because magnesium levels are low and nucleases are active. Stabilisation strategies such as oligolysine–polyethylene glycol coatings, lipid encapsulation, silica mineralisation, peptide or protein wraps and staple crosslinking can extend stability from minutes to many hours or days.
Can DNA origami be mass-produced?
Yes, at least in research-scale and pilot-scale demonstrations. Biotechnological production of scaffold and staple strands has shown that DNA origami can be made in macroscopic quantities, moving the field beyond early microgram-scale reactions. Clinical-grade manufacturing remains an active engineering and regulatory challenge.
Further Reading
Nature Reviews Methods Primers, DNA origami - a primer
Chemical Reviews, DNA Origami: Scaffolds for Creating Higher Order Structures
Nature Reviews Methods Primers, DNA origami
Nature Reviews Materials, Functionalizing DNA origami to investigate and interact with biological systems
