Nanobodies: Structure, Production, Applications, and Therapeutic Potential
What Are Nanobodies?
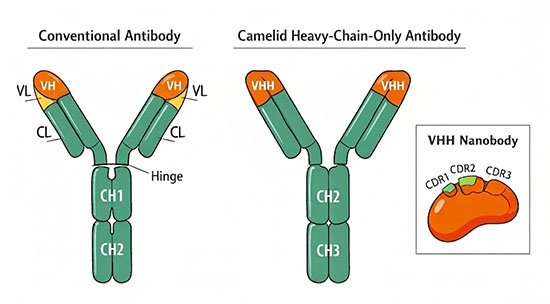
Nanobodies are single-domain antibody fragments derived from the variable region (VHH) of heavy-chain-only antibodies (HCAbs) naturally produced by members of the Camelidae family, including camels, llamas, and alpacas. With a molecular mass of approximately 12 to 15 kilodaltons and dimensions of roughly 2.5 nanometers in diameter by 4 nanometers in height, nanobodies are the smallest naturally occurring antibody fragments capable of binding an antigen with high affinity and specificity. Their formal designation in structural biology is VHH, though the term "nanobody," originally a registered trademark of the Belgian company Ablynx (now part of Sanofi), has become the standard name across research and clinical contexts.
The existence of heavy-chain-only antibodies was discovered in 1993 by a research group at Vrije Universiteit Brussel led by Raymond Hamers, who found that dromedary camel serum contained a substantial fraction of IgG-like molecules composed exclusively of heavy-chain dimers, lacking light chains entirely. This finding overturned the prevailing assumption that functional antibodies required both heavy and light chains for antigen recognition. Subsequent work by Serge Muyldermans and colleagues established methods for isolating and expressing the single VHH domain as a recombinant protein, laying the foundation for the nanobody field. Heavy-chain-only antibodies constitute approximately 60 to 80% of total serum IgG in dromedary and Bactrian camels and 25 to 50% in llamas and alpacas. A structurally analogous single-domain antibody, called VNAR, has also been identified in cartilaginous fish such as sharks, representing an independent evolutionary convergence on the same minimal antigen-binding scaffold.
Structure and Biophysical Properties
A nanobody consists of a single immunoglobulin variable domain folded into the canonical immunoglobulin β-sandwich structure: two β-sheets packed against each other and stabilized by a conserved intradomain disulfide bond. Like conventional VH domains, a nanobody contains four framework regions (FR1 to FR4) that provide the structural scaffold and three complementarity-determining regions (CDR1, CDR2, and CDR3) that form the antigen-binding surface, or paratope. The binding specificity and affinity of a nanobody are determined primarily by its CDR3 loop, which in camelid VHH domains tends to be longer than in human or mouse VH domains (often 15 to 20 residues compared to a typical 10 to 15) and can form extended finger-like projections that insert into cavities, enzyme active sites, and receptor clefts inaccessible to conventional flat or concave antibody paratopes.
A key structural adaptation that permits VHH domains to function without a light-chain partner is the substitution of several hydrophobic residues in FR2, specifically at positions 37, 44, 45, and 47 (Kabat numbering), with smaller, more hydrophilic amino acids. In conventional antibodies, these positions form the hydrophobic interface between VH and VL; in nanobodies, the substitution eliminates the need for VL pairing and simultaneously enhances solubility. Many VHH domains also contain an additional disulfide bond linking CDR3 to CDR1 or FR2, which stabilizes the elongated CDR3 loop and contributes to the high thermostability of nanobodies. Nanobodies commonly retain binding activity after incubation at 60 to 80 °C and can tolerate pH extremes (pH 3 to 9), chaotropic agents, and proteolytic exposure conditions that would denature conventional antibody fragments.
The small size and single-domain architecture of nanobodies confer several pharmacologically relevant properties. Their rapid renal clearance (serum half-life of approximately 1 to 2 hours in humans for a monomeric unmodified nanobody) is advantageous for molecular imaging, where fast blood clearance increases the signal-to-noise ratio, but is a limitation for therapeutic applications requiring sustained systemic exposure. Nanobodies achieve typical binding affinities in the low-nanomolar to picomolar range, with on-rates (kon) of 105 to 106 M−1s−1. Their high sequence homology to the human VH3 family contributes to intrinsically low immunogenicity, a feature supported by clinical data from approved nanobody-based drugs.
How Are Nanobodies Produced?
Antigen-specific nanobodies are most commonly obtained from immune libraries generated by immunizing a camelid, typically a llama or alpaca, with the target antigen over a course of four to eight weeks. After immunization, peripheral blood lymphocytes are harvested, and their mRNA is extracted and reverse-transcribed into cDNA. A two-step polymerase chain reaction (PCR) amplification strategy then selectively isolates VHH-encoding sequences from HCAb transcripts: a first PCR amplifies from the leader sequence to the CH2 domain, and the resulting amplicons are separated by agarose gel electrophoresis to distinguish the shorter HCAb-derived fragments (lacking CH1) from full-length conventional antibody heavy chains. A second, nested PCR amplifies the VHH coding region, which is then cloned into a phage display vector to create a library of typically 106 to 108 independent clones.
Phage display remains the dominant selection platform: each VHH is expressed as a fusion with the M13 phage coat protein pIII, and antigen-specific clones are enriched through iterative rounds of biopanning against the target. After two to three rounds of selection, individual clones are screened by ELISA, sequenced, and characterized for affinity and specificity. Alternative display technologies, including yeast surface display, bacterial display, and ribosome display, have expanded the toolkit, offering advantages such as quantitative affinity sorting by flow cytometry or entirely in vitro selection.
For targets where animal immunization is impractical, such as highly conserved self-antigens, toxic proteins, or conformational states that are difficult to present as immunogens, nanobodies can instead be selected from naïve or synthetic libraries. Naïve libraries are constructed from the VHH repertoire of non-immunized camelids and can yield binders against diverse targets without repeated immunization campaigns. Synthetic libraries are built entirely in vitro by grafting randomized CDR sequences onto a stable VHH scaffold, enabling complete control over library diversity and composition. A recombinant nanobody, once identified, is produced in microbial expression systems, most commonly E. coli or Pichia pastoris, at high yields and low cost, since the single-domain format requires no disulfide-mediated chain pairing and folds efficiently in bacterial periplasm or yeast secretory pathways.
Nanobodies Compared to Conventional Antibody Formats
The advantages and trade-offs of nanobodies relative to conventional monoclonal antibodies and other antibody fragments become clear when compared side by side. Conventional IgG antibodies offer bivalent binding, long serum half-life, and Fc-mediated effector functions. Nanobodies sacrifice these in monomeric form but gain deep tissue penetration, access to cryptic epitopes, extreme stability, and inexpensive microbial production.
| Feature | Conventional IgG | scFv | Nanobody (VHH) |
|---|---|---|---|
| Size | ~150 kDa | ~25 to 30 kDa | ~12 to 15 kDa |
| Serum half-life | ~21 days | Minutes to hours | ~1 to 2 hours (unmodified) |
| Tissue penetration | Limited | Moderate | Excellent |
| Epitope access | Flat/concave surfaces | Flat/concave surfaces | Cavities, clefts, active sites |
| Thermostability | Moderate | Low to moderate | High (60 to 80 °C) |
| Production host | Mammalian cells | Mammalian or bacterial | Bacterial or yeast |
| Fc effector functions | Yes | No | No (unless Fc-fused) |
These trade-offs have driven extensive engineering efforts to extend nanobody serum half-life. The most common strategies include fusion to an albumin-binding nanobody (which hitchhikes on serum albumin for FcRn-mediated recycling), direct genetic fusion to an Fc domain, and PEGylation. Multivalent and multispecific constructs such as bispecific nanobodies, tandem nanobodies, and nanobody-Fc fusions can combine targeting of two or more antigens in a single molecule while retaining production in microbial systems.
Therapeutic Applications
The therapeutic potential of nanobodies was validated in 2018 with the European approval (and February 2019 FDA approval) of caplacizumab (Cablivi), the first nanobody-based drug to reach the market. Caplacizumab is a bivalent anti-von Willebrand factor nanobody developed by Ablynx for the treatment of acquired thrombotic thrombocytopenic purpura (aTTP), a rare and life-threatening blood-clotting disorder. In the pivotal HERCULES trial, caplacizumab in combination with plasma exchange and immunosuppressive therapy significantly reduced the time to platelet count normalization and lowered the rate of aTTP-related death or recurrence compared to plasma exchange and immunosuppression alone.
Since caplacizumab's approval, several other nanobody-based therapeutics have entered late-stage clinical development or received approval. Ciltacabtagene autoleucel (Carvykti), a chimeric antigen receptor T cell (gene therapy) product for relapsed or refractory multiple myeloma, uses two anti-BCMA nanobodies in its CAR construct to achieve bivalent binding to the target antigen. Envafolimab, an anti-PD-L1 nanobody-Fc fusion, has been approved in China for microsatellite instability-high solid tumors. Ozoralizumab, a trivalent bispecific nanobody targeting TNF-α with an albumin-binding arm for half-life extension, received approval in Japan in 2022 for rheumatoid arthritis. Sonelokimab, a trivalent bispecific nanobody targeting both IL-17A and IL-17F, has demonstrated efficacy in phase 2b trials for psoriasis. These approvals and advanced clinical candidates span oncology, autoimmune disease, and hematology, reflecting the versatility of the nanobody platform.
In oncology, nanobodies are being explored across multiple therapeutic modalities. Nanobody-drug conjugates (NDCs) exploit the superior tumor penetration of nanobodies to deliver cytotoxic payloads directly to tumor cells. Bispecific nanobody constructs that simultaneously engage a tumor antigen and an immune effector cell (such as a T cell via CD3) are in development as alternatives to conventional bispecific antibodies. Nanobodies targeting immune checkpoints such as PD-L1, CTLA-4, and TIGIT are under investigation as next-generation biopharmaceuticals, and nanobody-based CAR-T constructs offer design simplicity over scFv-based CARs.
Applications in Diagnostics and Imaging
The rapid renal clearance that limits nanobody utility in sustained therapeutics becomes an asset in molecular imaging, where fast blood-pool clearance generates high contrast between target tissue and background within hours of injection. Nanobodies labeled with radionuclides such as 99mTc, 68Ga, and 18F are being developed as tracers for positron emission tomography (PET) and single-photon emission computed tomography (SPECT) imaging of tumors. Anti-HER2 nanobodies, for example, have demonstrated high-contrast tumor imaging in preclinical models and early clinical studies with same-day image acquisition, a significant workflow improvement over conventional antibody-based radiotracers that require 3 to 7 days for adequate contrast.
Beyond nuclear medicine, nanobodies serve as versatile detection reagents in in vitro diagnostics. Their stability under harsh conditions, including desiccation, elevated temperatures, and exposure to detergents, makes them robust components of lateral flow assays, biosensors, and immunoaffinity chromatography platforms. Nanobodies fused to reporter enzymes or fluorescent proteins can be produced as single fusion proteins in bacteria, eliminating the need for chemical conjugation steps. Anti-idiotypic nanobodies (nanobodies that bind other antibodies) are used as calibrators and toxin-free substitutes in food safety and environmental monitoring assays.
Nanobodies as Research Tools
In structural biology, nanobodies have become indispensable tools for stabilizing transient protein conformations and facilitating crystallization. G protein-coupled receptors (GPCRs), which are notoriously difficult to crystallize in their active states, have been solved in complex with conformationally selective nanobodies that lock the receptor into a defined conformation. The 2012 crystal structure of the β2-adrenergic receptor in its active state, work that contributed to Brian Kobilka's Nobel Prize in Chemistry, was enabled by a nanobody that stabilized the receptor's active conformation. Nanobodies are similarly used in cryo-electron microscopy as fiducial markers and conformational stabilizers.
Inside living cells, nanobodies function as intracellular binders, or intrabodies, that can modulate protein function, track protein localization, or recruit effector domains to specific targets. Chromobodies, fusions of a nanobody with a fluorescent protein, allow real-time visualization of endogenous proteins in living cells. The GFP-binding nanobody, one of the most widely used tools in cell biology, enables immunoprecipitation and subcellular protein redirection in GFP-tagged cell lines. Nanobody-based targeted protein degradation approaches exploit the modular format to direct specific proteins for proteasomal destruction.
Limitations and Challenges
The most significant limitation of monomeric nanobodies for therapeutic use is their short serum half-life. Rapid renal filtration of a 15 kDa protein means that unmodified nanobodies are cleared within hours, requiring either frequent dosing or half-life extension engineering. While albumin-binding fusions, Fc fusions, and PEGylation can extend half-life to days or weeks, each approach adds molecular complexity and may alter biodistribution. Balancing half-life extension with the retention of favorable tissue penetration remains an active engineering challenge.
The absence of an Fc domain in monomeric nanobodies means they lack the ability to recruit immune effector mechanisms such as antibody-dependent cellular cytotoxicity and complement activation. For applications where these functions are needed, nanobody-Fc fusion proteins restore effector activity but at the cost of increased size and the need for mammalian cell production to ensure proper glycosylation. The dependence on camelid immunization for high-quality immune libraries introduces logistical constraints, though synthetic and naïve libraries are increasingly viable alternatives.
From a regulatory perspective, nanobody-based therapeutics do not fit neatly into established monoclonal antibody frameworks. Their novel format and diverse engineering strategies (bispecific, trispecific, drug-conjugated) require case-by-case evaluation of pharmacokinetics, immunogenicity, and manufacturing consistency. As more nanobody products progress through clinical development, regulatory pathways are becoming better defined, but the field is still establishing the analytical standards that decades of monoclonal antibody experience have produced for conventional biologics.
Future Perspectives
Artificial intelligence and machine learning are beginning to transform nanobody discovery and optimization. Computational tools such as AlphaFold2 and nanobody-specific deep learning models (NanoNet, NanoBodyBuilder2) now predict VHH three-dimensional structures from sequence data with increasing accuracy, enabling in silico screening and rational design of CDR3 loops. AI-guided humanization algorithms can identify the minimal set of framework substitutions needed to reduce immunogenicity risk while preserving binding function, accelerating the transition from camelid-derived leads to clinical candidates.
The integration of nanobody technology with emerging therapeutic platforms is expanding the functional scope of these miniaturized binders. Nanobodies are being incorporated into targeted protein degradation systems, including proteolysis-targeting chimeras (PROTACs) and lysosome-targeting chimeras (LYTACs), where their binding properties direct specific proteins for degradation. In personalized medicine, multiplexed nanobody panels targeting different tumor-associated antigens could enable patient-specific diagnostic and therapeutic cocktails. As production costs continue to fall, nanobody stability under ambient conditions also positions them as components of point-of-care diagnostics for resource-limited settings, where cold-chain-independent reagents offer a critical practical advantage.
Frequently Asked Questions
What animals are used to produce nanobodies? Nanobodies are derived from the heavy-chain-only antibodies naturally found in members of the Camelidae family, which includes dromedary camels, Bactrian camels, llamas, alpacas, vicuñas, and guanacos. Llamas and alpacas are the most commonly used species in laboratory settings because they are docile, easy to house, and mount strong immune responses upon immunization. Heavy-chain-only antibodies with a similar single-domain architecture have also been identified in cartilaginous fish such as sharks and rays, though camelid-derived nanobodies dominate current research and clinical development.
How long does it take to develop a nanobody against a new target? The timeline depends on whether an immune, naïve, or synthetic library is used. For immune libraries, the immunization of a camelid typically takes six to eight weeks, followed by library construction, phage display selection, and characterization, which together require an additional four to eight weeks. A lead nanobody candidate can therefore be identified in roughly three to four months. Naïve and synthetic libraries bypass the immunization step entirely, allowing selection to begin immediately, though the resulting binders may require additional affinity maturation to match the potency of immune library-derived nanobodies.
Can nanobodies cross the blood-brain barrier? Several studies have demonstrated that certain nanobodies can cross the blood-brain barrier (BBB), a property largely attributed to their small size and favorable biophysical characteristics. Some nanobodies have been engineered to use receptor-mediated transcytosis pathways to reach the central nervous system. This has generated considerable interest in using nanobodies for molecular imaging and therapy of neurological conditions, including neurodegenerative diseases and brain tumors, though clinical validation of BBB-crossing nanobody therapeutics is still at an early stage.
Are nanobodies immunogenic in humans? Nanobodies have a high degree of sequence homology to human VH domains, particularly to the human VH3 family, which contributes to inherently low immunogenicity. Clinical experience with approved nanobody-based drugs such as caplacizumab has shown that treatment-related immunogenicity is generally mild. When further risk reduction is needed, nanobodies can be humanized by substituting camelid-specific framework residues with their human equivalents, a process that typically preserves binding affinity and stability.
What is the difference between a nanobody and a single-chain variable fragment (scFv)? Both nanobodies and scFvs are small antibody fragments used for antigen binding, but they differ in origin and structure. An scFv is an engineered construct that links the VH and VL domains of a conventional antibody with a flexible peptide linker (~25 to 30 kDa). A nanobody is a naturally occurring single-domain fragment (~12 to 15 kDa) from a camelid heavy-chain-only antibody and requires no partner domain. Nanobodies tend to be more stable, more soluble, and less prone to aggregation than scFvs, and they can be produced more readily in microbial hosts.
Further Reading
Annual Review of Biochemistry, Nanobodies: Natural Single-Domain Antibodies
The FEBS Journal, A Guide to: Generation and Design of Nanobodies
Journal of Nanobiotechnology, Discovery of Nanobodies: A Comprehensive Review of Their Applications and Potential over the Past Five Years
Molecular & Cellular Proteomics, Nanobodies: From High-Throughput Identification to Therapeutic Development
