Plasmid: Structure, Function, and Role in Biotechnology and Medicine
What Is a Plasmid?
A plasmid is a small molecule of DNA that exists separately from a cell’s main chromosomes and replicates independently of them. Most plasmids are circular and double-stranded, although linear plasmids exist in some species. Plasmids are found primarily in bacteria, where they carry accessory genes – for antibiotic resistance, virulence, or unusual metabolic capabilities – that are not essential for basic survival but provide selective advantages under specific conditions.
The term “plasmid” was coined by Joshua Lederberg in 1952 to describe extrachromosomal genetic elements observed during studies of bacterial conjugation and antibiotic resistance. Their transformation into biotechnology tools dates to 1973, when Stanley Cohen and Herbert Boyer with their colleagues at Stanford and the University of California, San Francisco demonstrated that DNA fragments cut with restriction enzymes could be ligated into a bacterial plasmid and replicated inside Escherichia coli. This experiment marked the birth of recombinant DNA technology, and by 1982 the same plasmid-based approach had enabled the first commercial production of human insulin from genetically modified bacteria.
Plasmids range in size from about 1,000 base pairs to more than 1 million, and a single bacterium can carry several plasmids at different copy numbers. The Addgene plasmid repository alone distributes more than 170,000 unique plasmids deposited by nearly 7,000 laboratories and has shipped over 2 million samples since 2004. Plasmid DNA is also a regulated biopharmaceutical raw material manufactured at industrial scale for use in DNA vaccines and gene therapy production.
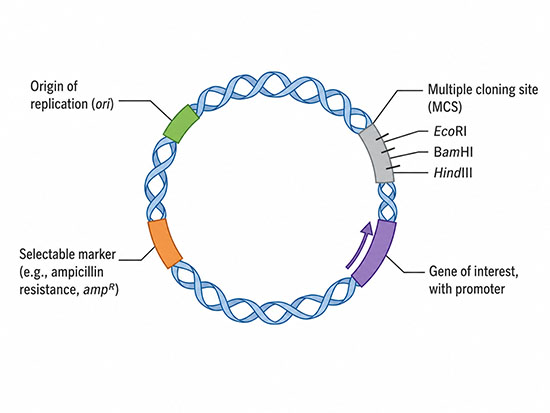
Structure and Key Features of Plasmids
Most plasmids are covalently closed circular DNA molecules that exist in a tightly twisted, supercoiled topology. Supercoiled plasmid is more compact than relaxed circular DNA and is preferred for laboratory and biopharmaceutical applications because it transfects host cells more efficiently and is the form recognized as the active biopharmaceutical species by regulators. Plasmid sizes span four orders of magnitude, from small cloning vectors of 2,000 to 10,000 base pairs to megaplasmids in some soil and rhizobial bacteria that exceed 1 million base pairs. The largest of these blur the line between plasmid and chromosome and are sometimes classified as “chromids,” a hybrid category.
A functional plasmid contains a small number of essential elements regardless of its purpose. The origin of replication (ori) is the sequence at which DNA polymerase initiates copying of the plasmid, and it determines both the host range – which bacterial species can maintain the plasmid – and the copy number per cell. A selectable marker, usually an antibiotic resistance gene, allows researchers to identify plasmid-containing cells by growing them on antibiotic-containing medium. A multiple cloning site (MCS), a short region with several unique restriction enzyme recognition sequences, provides convenient positions for inserting foreign DNA.
Plasmids designed for gene expression in a particular host carry additional elements appropriate to that host: a strong promoter, a ribosome-binding site for bacterial expression or a Kozak sequence for mammalian expression, and a transcription terminator or polyadenylation signal flanking the cloning site. Shuttle vectors carry two origins of replication so they can be propagated alternately in bacteria and in yeast or human cells, and expression vectors for proteins often include affinity tags such as polyhistidine sequences to simplify downstream purification.
Plasmid Replication and Inheritance
Plasmid replication is independent of chromosomal replication and is controlled by the ori sequence and a small number of plasmid-encoded proteins. Copy number varies widely with the type of ori. Stringently controlled plasmids such as the F factor are maintained at one or two copies per cell; the wild-type ColE1 plasmid sits at roughly 15–20 copies; and pUC-derived high-copy cloning vectors, which carry a mutation that disables ColE1 copy-number control, can reach 500 to 700 copies per cell. This high yield allows milligram quantities of plasmid DNA to be recovered from a single liter of bacterial culture.
Faithful inheritance of plasmids during cell division is not automatic for low-copy plasmids; if segregation were left to chance, daughter cells would frequently inherit no plasmid copies. Many natural plasmids therefore carry partition systems that actively distribute plasmid copies to opposite poles of the cell before division, and post-segregational killing systems – toxin–antitoxin modules in which a stable toxin kills any daughter cell that loses the plasmid – that selectively eliminate plasmid-free progeny. These maintenance mechanisms explain how costly antibiotic resistance plasmids can persist in bacterial populations even when the corresponding antibiotic is absent.
Plasmids in Nature: Antibiotic Resistance and Horizontal Gene Transfer
In nature, plasmids are central agents of bacterial adaptation. Conjugative plasmids encode the protein machinery – including a surface pilus and a relaxase enzyme – needed to copy themselves into a neighboring bacterium through direct cell-to-cell contact, a process called conjugation. Mobilizable plasmids can be transferred when conjugation machinery is supplied in trans by another plasmid in the same cell. Through conjugation, transformation, and transduction by bacteriophages, plasmids move between bacterial species and even between distantly related genera, redistributing accessory genes across diverse hosts in what is collectively termed horizontal gene transfer.
This mobility makes plasmids the most clinically important vehicle for the spread of antimicrobial resistance. Plasmids carry resistance determinants for nearly every class of clinically used antibiotic, and a single multidrug-resistance plasmid can confer resistance to several drug families simultaneously. The 2016 emergence of the plasmid-borne mcr-1 gene, which confers resistance to colistin – an antibiotic of last resort for infections by carbapenem-resistant Gram-negative bacteria – illustrated how rapidly plasmid-mediated resistance can spread between agricultural and clinical settings on a global scale, and motivated the integration of plasmid surveillance into the One Health framework that links human, animal, and environmental microbiology.
Plasmids also play a broader role in bacterial evolution that goes beyond simply ferrying resistance genes. Because plasmids are typically present at multiple copies per cell, they create islands of polyploidy within an otherwise haploid bacterial genome. This polyploidy can buffer the effects of new mutations, allow heterozygosity at single loci, and accelerate the evolution of plasmid-encoded genes relative to their chromosomal counterparts. New traits such as antibiotic resistance and virulence often appear first on plasmids and only later are captured by the chromosome.
Plasmids as Tools in Molecular Biology and Genetic Engineering
The same features that make plasmids successful in nature – compact replicons, selectable markers, and built-in transfer machinery – are precisely what make them useful in the laboratory. The Cohen–Boyer experiment of 1973 transformed plasmids from objects of microbial study into the foundational tool of genetic engineering and modern molecular biology. A typical cloning workflow cuts a fragment of interest and a plasmid vector with the same restriction enzymes, ligates them, and introduces the recombinant construct into E. coli. Plasmid-containing cells are then selected on antibiotic-containing medium and verified by DNA sequencing.
Beyond simple cloning, specialized plasmid vectors support a wide range of laboratory functions. Expression vectors carrying strong inducible promoters allow the production of recombinant proteins in bacterial, yeast, insect, or mammalian cells; this is the basis of industrial production of human insulin and growth hormone. Plasmids deliver the guide RNAs and Cas9 nuclease used in CRISPR-Cas9 genome editing experiments; they encode the antibody chains and selection markers used in monoclonal antibody display libraries; and they form the backbone for protein engineering work in directed evolution and rational design.
In synthetic biology, entire metabolic pathways comprising dozens of genes are assembled on a single large plasmid using techniques such as Gibson assembly and Golden Gate cloning, then introduced into bacteria, yeast, or plant cells to produce bioplastics, biofuels, terpenoids, or pharmaceutical precursors. Standardized plasmid backbones with well-characterized parts – promoters, ribosome binding sites, terminators – allow synthetic biologists to design genetic circuits the way electrical engineers design integrated circuits, and online repositories such as Addgene and the iGEM Registry of Standard Biological Parts make tens of thousands of these characterized plasmid components openly available to researchers worldwide.
Plasmids in Biopharmaceutical Manufacturing and Therapy
Plasmid DNA serves the biopharmaceutical industry in two distinct roles: as a raw material for the manufacture of viral vectors, and as a finished drug substance in its own right. The two roles share a manufacturing process – fermentation in E. coli followed by purification of supercoiled plasmid – but face different regulatory requirements. Adeno-associated virus (AAV) vector manufacturing for in vivo gene therapy requires co-transfection of three plasmids – encoding the viral rep and cap genes, adenoviral helper functions, and the therapeutic transgene – into HEK293 cells in a bioreactor. Lentiviral vector production for CAR-T cell therapy similarly relies on transient transfection of three or four plasmids into HEK293T cells.
Plasmid DNA can also be used directly as a biopharmaceutical. DNA vaccines consist of a plasmid encoding a pathogen antigen under the control of a mammalian promoter; once delivered into muscle or skin cells, the plasmid is transcribed in the nucleus and translated to produce the antigen, triggering an adaptive immune response without the use of live virus or recombinant protein. Because skeletal muscle fibers are post-mitotic, plasmid taken up after intramuscular injection can persist and continue expressing antigen for months. The first DNA vaccine for human use, ZyCoV-D against SARS-CoV-2, received emergency use authorization in India in August 2021.
Although DNA vaccines have proven less immunogenic than mRNA vaccines in humans, their stability at room temperature, simple manufacturing, and ease of redesign for new pathogens or tumor antigens continue to motivate clinical development for cancer immunotherapy, HIV, and diseases requiring durable cellular immunity. Active areas of optimization include codon-optimized antigen sequences, electroporation and needle-free jet delivery to improve cellular uptake, and co-expression of plasmid-encoded molecular adjuvants such as cytokines and chemokines. Several DNA-based therapies for veterinary indications, including a melanoma vaccine for dogs and growth-hormone-releasing-hormone plasmid products for swine, are already commercialized.
Frequently Asked Questions
What is the difference between a plasmid and a chromosome? Both plasmids and chromosomes are DNA molecules inside a cell, but they differ in size, content, and how they are inherited. The bacterial chromosome typically contains a few thousand essential genes and is present as a single copy that is replicated and segregated in tight coordination with cell division. A plasmid is much smaller, usually carries accessory genes that are useful but not essential for survival, and replicates independently of the chromosome. A single bacterium can carry multiple plasmids of different copy numbers, while it normally carries only one chromosome.
Are plasmids found in human cells? Naturally occurring plasmids are essentially absent from human cells and from most multicellular animals. Plasmids are characteristic of bacteria, archaea, and some single-celled eukaryotes such as yeast. Mitochondrial DNA, although small and circular, is part of the cell’s heritable genome rather than an accessory replicon and is not classified as a plasmid. When plasmid DNA enters a human cell during transfection or DNA vaccination, it is transcribed in the nucleus but is not stably maintained or replicated; in dividing cells it is gradually diluted and lost, while in long-lived post-mitotic cells such as skeletal muscle fibers it can persist and continue expressing antigen for months.
Why do plasmids carry antibiotic resistance genes in laboratory experiments? An antibiotic resistance gene on a research plasmid acts as a selectable marker, not as a clinical resistance trait. After researchers attempt to introduce a plasmid into bacteria, only a small fraction of the cells actually take it up. Plating the cells on medium containing the corresponding antibiotic kills the bacteria that lack the plasmid and allows only the plasmid-containing cells to grow into visible colonies. The most common laboratory markers are resistance to ampicillin, kanamycin, and chloramphenicol – antibiotics chosen for their effectiveness in laboratory strains rather than their clinical importance.
Can plasmids replicate outside a cell? Plasmids cannot self-replicate outside a living host cell. Although plasmids encode the proteins that initiate their own replication, they depend on the host’s DNA polymerase, nucleotide pool, and energy metabolism to actually copy themselves. Outside a cell, purified plasmid DNA is a stable molecule that can be stored for years at low temperature, but it remains inert until it is reintroduced into a compatible bacterial host or transfected into a eukaryotic cell capable of transcribing and translating the genes it carries.
How are plasmids different from viral vectors for gene delivery? Plasmids and viral vectors are both used to deliver foreign genes into cells, but they differ in efficiency, capacity, and integration behavior. Naked plasmid DNA enters cells inefficiently and is expressed transiently without integrating into the host genome, which is an advantage for safety but a limitation for durable expression. Viral vectors such as adeno-associated virus and lentivirus exploit natural viral entry mechanisms to achieve much higher transduction efficiency in human cells, and lentiviral vectors integrate into the host genome for long-term expression. Plasmids are typically used for laboratory cloning, DNA vaccines, and as raw materials for manufacturing viral vectors themselves.
Further Reading
Proceedings of the National Academy of Sciences, Construction of Biologically Functional Bacterial Plasmids In Vitro
Nature Reviews Microbiology, Beyond Horizontal Gene Transfer: The Role of Plasmids in Bacterial Evolution
Nature Reviews Microbiology, Plasmids, a Molecular Cornerstone of Antimicrobial Resistance in the One Health Era
Microbiology Spectrum, Plasmid Biopharmaceuticals
Microbial Biotechnology, New Insights for the Development of Efficient DNA Vaccines
