Restriction Enzyme: Definition, Types, Sticky Ends, and Uses
Definition: A restriction enzyme is a DNA-cutting protein, found mainly in bacteria and archaea, that recognizes a specific short DNA sequence and cleaves the double helix at or near that site. By producing predictable, reproducible DNA fragments, restriction enzymes act as the molecular scissors that made recombinant DNA technology and modern molecular cloning possible.
What Is a Restriction Enzyme?
A restriction enzyme, also called a restriction endonuclease, is an enzyme that scans double-stranded DNA, binds a defined short sequence known as a recognition site, and cleaves the sugar-phosphate backbone of both strands. Recognition sites are typically four to eight base pairs long and are often palindromic, reading the same on each strand in the 5′-to-3′ direction. These enzymes are part of microbial defense systems: bacteria and archaea use them to “restrict” invading viral DNA by cutting it into pieces, while protecting their own genome by chemically tagging the same sequences. This paired cutting-and-protecting machinery is called a restriction-modification system.
Restriction enzymes were discovered through studies of why some bacteriophages grew poorly on certain bacterial strains. The work earned Werner Arber, Hamilton Smith, and Daniel Nathans the 1978 Nobel Prize in Physiology or Medicine. The first site-specific restriction enzyme was isolated from the bacterium Haemophilus influenzae in 1970, and within a few years these proteins had become indispensable laboratory reagents. Thousands of restriction enzymes with several hundred distinct recognition specificities have since been characterized and catalogued in the curated REBASE database.
Their importance comes from a single property: a given enzyme always cuts at the same sequence. DNA from any organism can therefore be reduced to a defined, repeatable set of fragments rather than a random smear, turning an unreadable chromosome into a collection of manageable, recombinable pieces. That reproducibility is the foundation on which gene cloning, physical mapping of DNA, and much of the modern biotechnology industry were built.
At a glance:
- What it is: a bacterial or archaeal enzyme that cuts double-stranded DNA at a specific sequence
- Recognition site: usually 4–8 base pairs, frequently palindromic
- Biological role: defense against bacteriophages (restriction-modification system)
- Cofactor: Type II enzymes require Mg2+
- Cut products: sticky (cohesive) ends or blunt ends
- Main use: molecular cloning, recombinant DNA, DNA mapping, and DNA assembly
In this article: how restriction enzymes work; the main enzyme types; sticky versus blunt ends; how restriction enzymes are used in cloning and DNA assembly; and how they differ from programmable genome editors such as CRISPR-Cas9.
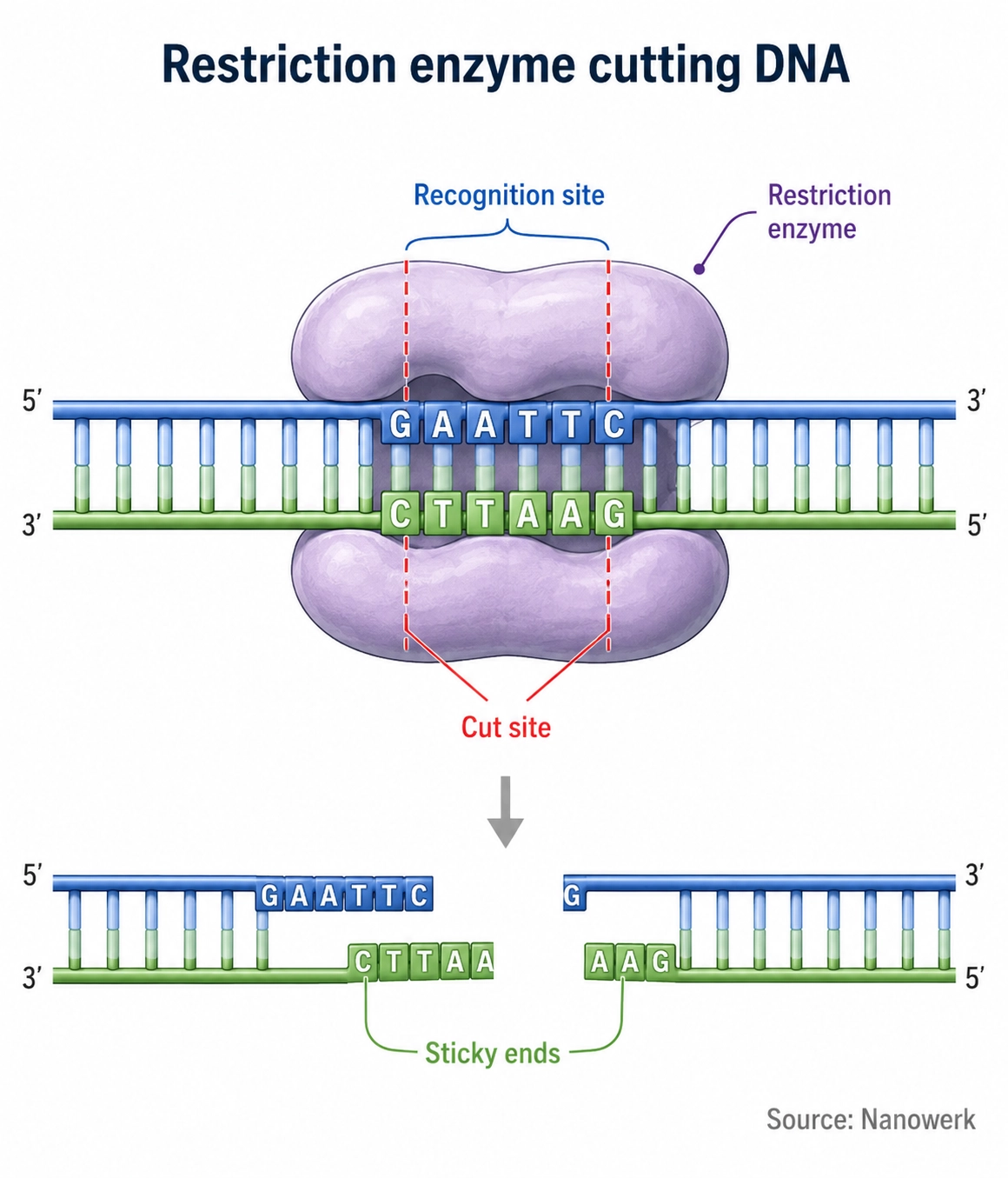
How Do Restriction Enzymes Work?
A restriction enzyme finds its target by sampling the DNA, often sliding or hopping along the helix until it encounters its recognition sequence. Recognition is highly specific: the protein contacts the edges of the bases, mainly in the major groove of the double helix, forming a precise network of hydrogen bonds and other contacts that fit only the correct sequence. Most of the enzymes used in the laboratory are symmetric dimers that bind a palindromic site so that each half of the protein reads one strand, which explains why so many recognition sequences are palindromes.
Once correctly bound, the enzyme catalyzes hydrolysis of the phosphodiester bonds in the DNA backbone, breaking one bond on each strand. For the common Type II enzymes used in cloning, this reaction requires magnesium ions, Mg2+, as a cofactor, and it leaves a phosphate group on one cut end and a hydroxyl group on the other – the chemistry that allows the fragments to be rejoined later by a DNA ligase. The enzyme typically releases the cut DNA and moves on, having converted one long molecule into shorter, defined fragments.
Specificity is the defining feature. A change of even a single base within the recognition site is usually enough to prevent cutting, which is also how the host bacterium protects itself: a companion methyltransferase adds a methyl group to a base within each recognition site in the cell’s own DNA, making it invisible to the matching restriction enzyme while leaving unmethylated phage DNA vulnerable. Under non-ideal reaction conditions, some enzymes show relaxed cutting at near-match sequences, a phenomenon known as star activity that modern high-fidelity variants are engineered to suppress.
Main Types of Restriction Enzymes
Restriction enzymes are commonly grouped into Types I, II, III, and IV according to their subunit structure, cofactors, methylation behavior, and where they cut relative to the recognition site. Type II enzymes are the most important laboratory tools because they cut at predictable positions. Type IIS enzymes are an important subgroup of Type II enzymes: they recognize an asymmetric sequence but cut a short distance outside it, which makes them especially useful for scarless DNA assembly.
| Type | Where it cuts | Cofactors | Main significance |
|---|---|---|---|
| Type I | Far from the recognition site, at a variable position | ATP, S-adenosylmethionine, Mg2+ | Large restriction-modification machines; important biologically but not useful for routine cloning |
| Type II | At or very close to the recognition site, usually at a predictable position | Usually Mg2+ | Standard laboratory tools for cloning, restriction mapping, plasmid verification, and DNA assembly |
| Type IIS (Type II subgroup) | A fixed, short distance outside an asymmetric recognition site | Mg2+ | Produces custom overhangs for scarless, multi-fragment assembly methods such as Golden Gate cloning |
| Type III | A short distance away from the recognition site, often requiring two oppositely oriented sites | ATP, Mg2+ | More complex restriction-modification systems; mainly of biological interest |
| Type IV | Modified DNA, especially methylated or otherwise chemically marked DNA | Varies; often nucleotide-dependent | Targets DNA carrying modification patterns rather than protecting those sites; specialized research interest |
The non-obvious point is that the most biologically elaborate enzymes are usually the least convenient laboratory cutters. Type I and Type III systems are remarkable molecular machines, but their cleavage behavior is too distant or context-dependent for simple excision of defined fragments. Many commonly used Type II enzymes are simpler proteins with separate methyltransferases, and they became the workhorses of molecular biology because they cut reproducibly. Type IIS enzymes extend that usefulness by cutting outside their recognition sequence, allowing the recognition site to be removed during assembly.
Recognition Sites, Sticky Ends, and Blunt Ends
Many Type II recognition sites are palindromes. A classic example is the sequence GAATTC, cut by the enzyme EcoRI. Because the bottom strand read in its own 5′-to-3′ direction is also GAATTC, a symmetric enzyme can engage both strands identically. Where the enzyme breaks the backbone within that site determines the shape of the resulting DNA ends, and that shape has large practical consequences.
When an enzyme cuts the two strands at staggered positions, it leaves short single-stranded overhangs called sticky or cohesive ends. EcoRI, for instance, produces ends with a single-stranded AATT overhang. When an enzyme cuts both strands at the same position, it produces flush, double-stranded blunt ends. Sticky ends are valuable because the overhang from one fragment can base-pair with a complementary overhang on any other fragment cut by the same enzyme, holding the pieces together so a ligase can seal them. This ability to make DNA from completely different sources join through matching overhangs is the conceptual heart of recombinant DNA.
Restriction Enzymes in Molecular Cloning and Recombinant DNA
In the early 1970s, researchers showed that a restriction enzyme could cut a circular plasmid and a piece of foreign DNA so that the two could be stitched together with DNA ligase and propagated inside Escherichia coli. That experiment created the first recombinant DNA molecules and launched genetic engineering as a practical discipline. The logic remains standard today: cut a vector and an insert with the same enzyme or enzymes, mix them so their compatible ends anneal, seal the junctions with ligase, and introduce the construct into cells.
Cloning plasmids were redesigned to exploit this. They carry a short stretch of DNA called a multiple cloning site, or polylinker, that packs many unique restriction sites into a small region. Cutting a plasmid with two different enzymes lets a gene be inserted in a single, defined orientation, a technique called directional cloning. The same enzymes underpin restriction mapping, in which the pattern of fragment sizes produced by various enzymes is used to deduce the arrangement of sites along a DNA molecule, and they remain a routine part of molecular cloning workflows across molecular biology.
Applications in Research, Diagnostics, and Synthetic Biology
Beyond cloning, restriction enzymes have a long history in analysis and diagnostics. Restriction fragment length polymorphism analysis, which detects sequence differences between individuals as changes in fragment sizes, was an early workhorse of genetic mapping and DNA fingerprinting before sequencing became cheap. Restriction digestion is still used to verify that a plasmid has the expected structure, to screen bacterial colonies for correct inserts, and to prepare DNA fragments for analysis or for DNA sequencing library preparation.
The Type IIS enzymes, which cut a fixed distance outside their recognition sequence, enabled a newer generation of assembly methods. Because the cut leaves a custom-designed overhang that does not contain the recognition site, many fragments can be digested and ligated together in a single tube to build large constructs without unwanted scars – the principle behind Golden Gate assembly and related techniques now central to synthetic biology. These methods are used to construct multi-gene pathways, build viral vectors for gene therapy research, and assemble guide-RNA and editing constructs, linking a 1970s discovery to the most current genome engineering. Computational tools and databases such as REBASE, part of the broader toolkit of bioinformatics, catalogue enzyme specificities and help researchers design these strategies.
Restriction Enzymes and the Move to Programmable Editing
Restriction enzymes have one fundamental limitation as editing tools: their recognition sequences are fixed by evolution and cannot be reprogrammed at will. A four-to-eight base-pair site also usually occurs many times across a large genome, so a restriction enzyme generally cannot be aimed at one unique chromosomal location the way precise genome surgery requires. These constraints drove the development of programmable nucleases – first zinc-finger nucleases, then TALENs, and most prominently CRISPR-Cas9 – whose specificity can be redirected to almost any chosen sequence.
This does not make restriction enzymes obsolete. For cutting and reassembling DNA in the test tube – the daily business of cloning, plasmid building, and library preparation – their speed, reliability, and low cost continue to make them standard tools, and the programmable editors are themselves often built using restriction-based assembly. The two technologies occupy different niches: restriction enzymes for manipulating DNA in vitro, programmable nucleases for gene editing inside living cells. Together they continue to support a large part of biotechnology.
Frequently Asked Questions
How are restriction enzymes named? Restriction enzyme names are derived from the organism they were isolated from. The first letter comes from the genus and the next two from the species, followed by a strain designation and a Roman numeral indicating the order of discovery in that organism. EcoRI, for example, was the first enzyme characterized from Escherichia coli strain RY13, and HindIII is the third from Haemophilus influenzae serotype d.
What is the difference between an endonuclease and an exonuclease? An endonuclease cuts a nucleic acid chain internally, between two nucleotides somewhere along the strand. An exonuclease instead removes nucleotides one at a time from a free end of the strand. Restriction enzymes are endonucleases because they cleave the DNA backbone at internal recognition sites rather than chewing inward from the ends.
Can restriction enzymes cut RNA? Classical restriction enzymes act on double-stranded DNA, not RNA. Their recognition and catalysis evolved for the DNA double helix, and they generally do not cleave single-stranded nucleic acids or RNA-DNA hybrids efficiently. Cutting RNA at defined sequences requires different tools, such as ribozymes or RNA-targeting CRISPR systems.
What is star activity in restriction enzymes? Star activity is off-target cutting by a restriction enzyme under non-ideal reaction conditions. It can happen when glycerol level, enzyme concentration, salt, pH, or incubation time falls outside the recommended range. Modern high-fidelity enzyme variants are engineered to minimize this behavior.
Are restriction enzymes still used now that CRISPR exists? Yes. CRISPR-Cas9 is programmable and is preferred for editing a chosen site inside a living genome, but restriction enzymes remain standard tools for cutting and reassembling DNA in the test tube. Cloning, plasmid construction, restriction mapping, and modern DNA assembly methods still depend heavily on them, so the two technologies are complementary rather than competing.
Further Reading
Proceedings of the National Academy of Sciences, How Restriction Enzymes Became the Workhorses of Molecular Biology
Nucleic Acids Research, Highlights of the DNA Cutters: A Short History of the Restriction Enzymes
Nucleic Acids Research, Type II Restriction Endonucleases — A Historical Perspective and More
Nucleic Acids Research, REBASE: A Database for DNA Restriction and Modification: Enzymes, Genes and Genomes
