Molecular Cloning: Definition, Workflow, Methods, and Uses
Definition: Molecular cloning is a set of laboratory techniques for making many identical copies of a defined DNA fragment by joining it to a self-replicating carrier molecule, called a vector, and propagating that construct inside a living host cell.
What Is Molecular Cloning?
Molecular cloning is the process of isolating a specific piece of DNA – most often a single gene – attaching it to a carrier DNA molecule that can replicate on its own, and introducing the combined molecule into a host cell that copies it faithfully every time it divides. The product is a population of cells that all carry the same engineered DNA, and from which large amounts of that exact sequence can be recovered. Because the inserted fragment and its carrier come from different sources, the resulting molecule is a form of recombinant DNA.
The term is often confused with the cloning of whole organisms, but the two refer to very different laboratory practices. Molecular cloning copies a defined DNA sequence, not a whole animal; although selected host cells form clonal colonies, the goal is to recover or use the engineered DNA construct. The shared word reflects only that both involve making genetically identical copies. The single fragment carried by one bacterial cell can become billions of identical copies in an overnight culture, and a typical plasmid vector is only about 3,000 to 10,000 base pairs long. Introduced in the early 1970s, molecular cloning is one of the foundational techniques of genetic engineering and modern molecular biology.
At a glance:
- Core idea: copy one defined DNA fragment by replicating it inside a living host
- Key parts: an insert (the DNA of interest), a vector (the carrier), a host cell
- Classic workflow: cut → ligate → transform → select → screen
- Workhorse host: Escherichia coli
- Origin: first recombinant plasmids constructed in 1972–1973
- Modern shift: overlap-based and type IIS assembly methods such as Gibson Assembly and Golden Gate alongside classical restriction–ligation
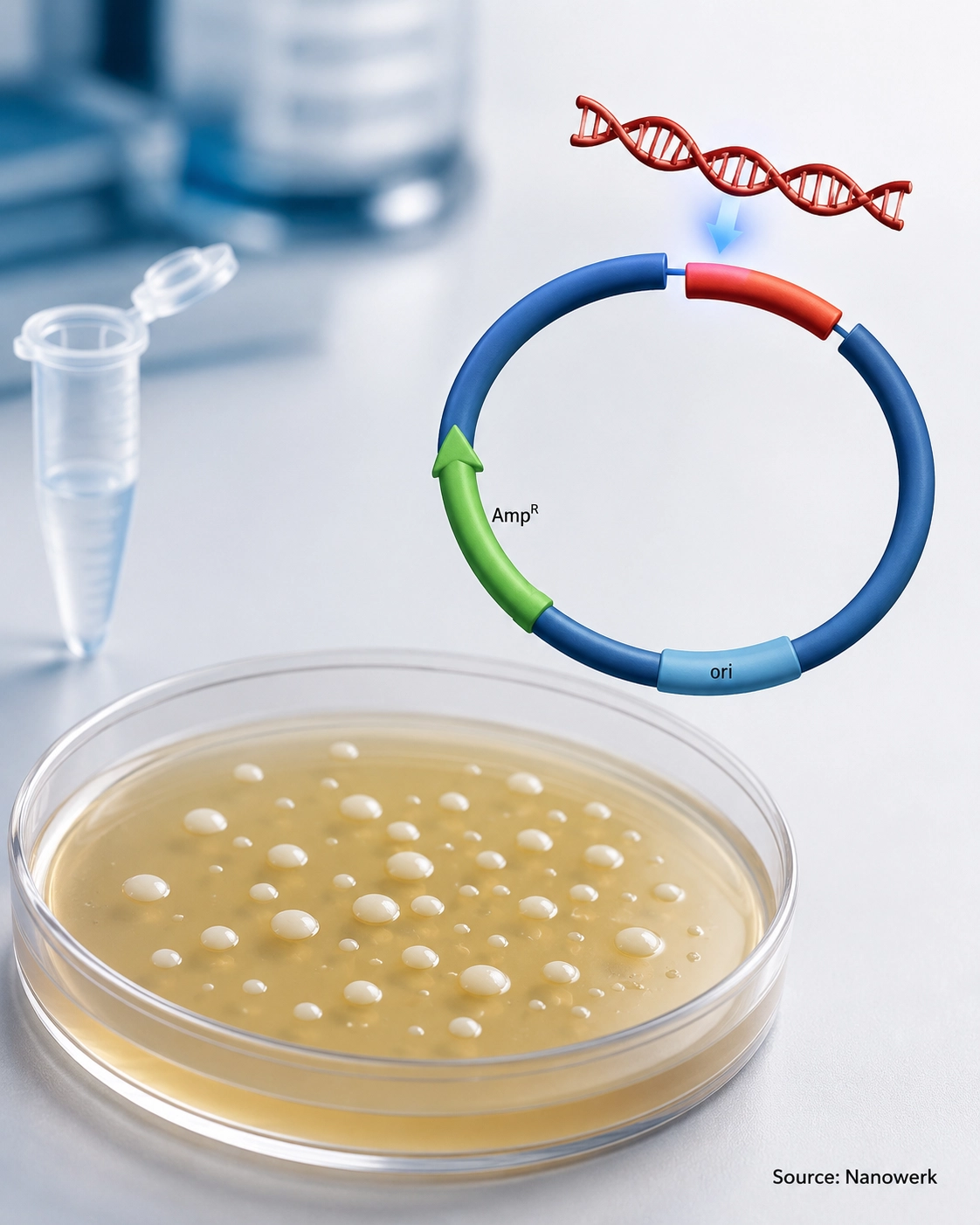
How Does Molecular Cloning Work?
The classical workflow has five conceptual stages: obtain the DNA fragment to be cloned, cut both the fragment and the vector so they can be joined, ligate them into a single recombinant molecule, introduce that molecule into a host cell, and finally select and verify the cells that carry the correct construct. Each stage relies on enzymes that act on DNA with high specificity, which is what makes the process reliable enough to be a routine laboratory tool.
The fragment of interest can be obtained in several ways. It may be cut directly from genomic DNA, copied from a messenger RNA template as complementary DNA, amplified by the polymerase chain reaction, or, increasingly, ordered as a chemically synthesized oligonucleotide or synthetic gene. PCR is especially common because it lets a researcher add tailored sequences to the ends of the fragment that make later joining steps easier.
Cutting and joining are the chemical heart of classical cloning. Restriction endonucleases are enzymes, originally discovered in bacteria, that recognize short, often palindromic DNA sequences and cut the double helix at or near those sites. Many leave short single-stranded overhangs, called sticky ends, that pair with complementary overhangs on a vector cut with the same enzyme; others leave blunt ends. DNA ligase then forms the covalent bonds that seal the fragment and vector into one continuous circular molecule, the recombinant construct.
The construct must next enter a host cell to be copied. In bacterial transformation, chemically treated E. coli cells take up plasmid DNA after a brief heat shock, or DNA is driven across the membrane by a short electrical pulse in electroporation. Once inside, the plasmid uses its own origin of replication and the host's machinery to copy itself; under selective conditions, most daughter cells maintain the construct as the population grows.
Because only a minority of cells take up a correctly assembled plasmid, the final stage is selection and screening. Vectors carry a selectable marker, typically an antibiotic-resistance gene, so cells that have taken up a plasmid carrying the marker are able to grow on a selective plate. Further checks – blue/white colony screening, colony PCR, restriction digestion, and confirmatory DNA sequencing – confirm that a colony carries the intended insert in the correct orientation before it is used or stored.
The Cloning Toolkit: Vectors, Enzymes, and Host Cells
A vector is any DNA molecule that can carry an insert and replicate inside a host. Plasmids, small circular DNA molecules from bacteria, are the most common. Standard plasmid vectors are best suited to small-to-moderate inserts, often up to about 10 kilobases in routine work, although specialized plasmids can carry larger fragments. Larger payloads need other vectors: bacteriophage and cosmid vectors hold tens of kilobases, bacterial artificial chromosomes (BACs) commonly carry hundreds of kilobases, and yeast artificial chromosomes (YACs) can extend into megabase-scale inserts, which made artificial chromosome vectors central to genome projects. Viral vectors are used when the goal is to deliver DNA into mammalian cells rather than simply amplify it.
Vectors also differ by purpose. A cloning vector is built mainly to maintain and amplify an insert, whereas an expression vector adds regulatory elements – promoters and other control sequences such as ribosome-binding sites in bacteria, Kozak sequences in eukaryotes, terminators, or polyadenylation signals – that drive gene expression so the cloned gene is transcribed and translated into protein. The enzyme toolkit beyond restriction endonucleases and ligases includes DNA polymerases for amplification and gap filling, exonucleases that chew back DNA ends to create overlaps, and site-specific recombinases used by some modern systems.
Host choice depends on the application. E. coli is the universal workhorse for amplifying and storing DNA because it grows fast and takes up plasmids readily. Yeast supports the assembly of very large constructs through its efficient homologous recombination. Mammalian and insect cells grown in cell culture are used when correctly folded, modified human proteins are needed, since bacteria cannot perform many eukaryotic protein modifications.
Classical Cloning Versus Modern DNA Assembly
Classical restriction–ligation cloning was transformative but has structural limits: it depends on suitable restriction sites being present in convenient places and absent from the insert, it usually leaves small unwanted "scar" sequences at the junctions, and joining several fragments at once is difficult. These constraints drove the development of newer methods that are largely sequence-independent and can stitch many fragments together in a single reaction, a capability that became essential as synthetic biology moved toward building large, multi-part constructs.
The main families now in routine use trade off scarring, the number of fragments they can join, site dependency, and cost. The table below summarizes how the dominant approaches compare.
| Method | Joining principle | Multi-fragment capacity | Leaves a scar? | Typical use |
|---|---|---|---|---|
| Restriction–ligation | Restriction-enzyme cut plus DNA ligase | Low (1–2 inserts) | Usually yes | Simple single-gene constructs, teaching |
| TA / TOPO cloning | PCR-product overhangs or topoisomerase | Low | Often yes | Fast capture of PCR products |
| Gateway recombination | Site-specific recombinases | Moderate | Yes (att sites) | Transferring inserts across vector sets |
| Gibson Assembly | Exonuclease, polymerase and ligase on overlaps | High | No when overlaps are designed for seamless junctions | Multi-fragment and large constructs |
| Golden Gate | Type IIS enzymes cutting outside their site | High | Usually seamless; junctions are user-designed | Modular, standardized part assembly |
The non-obvious trade-off is that no single method is best for everything. Restriction–ligation remains the simplest and cheapest route for a one-fragment construct and is still the standard teaching method, despite its scars and site dependence. Overlap-based and type IIS methods such as Gibson Assembly and Golden Gate excel at joining many parts precisely, but they require careful overlap, junction, or overhang design and assume a known sequence, so they are less convenient for capturing an uncharacterized fragment quickly. In practice, laboratories keep several techniques available and match the method to the construct rather than standardizing on one.
What Is Molecular Cloning Used For?
One of the most economically significant applications is the production of recombinant proteins. By cloning a human gene into an expression vector and growing the host at scale, manufacturers produce therapeutic proteins that were once scarce or unavailable; recombinant human insulin, first marketed in the early 1980s, was the prototype, and the same logic underlies biopharmaceuticals from clotting factors to antibodies. Cloning is also the routine way to obtain pure protein for structural and functional study, and to build the gene libraries used to survey genomes and transcriptomes.
Beyond protein production, molecular cloning is the construction step behind much of modern biotechnology. Building the plasmids and viral vectors used in gene therapy requires cloning; assembling the promoter, nuclease, and guide-RNA cassettes used in CRISPR-Cas9 gene-editing tools is itself a cloning task; and the engineered pathways and circuits of synthetic biology are assembled from cloned parts. Cloning libraries of gene variants likewise feeds directed evolution and protein engineering, and it remains central across biotechnology research and manufacturing.
- Producing recombinant proteins such as insulin, enzymes, clotting factors, and antibodies
- Building plasmids and viral vectors for gene therapy and gene-editing workflows
- Creating gene, cDNA, and variant libraries for screening and discovery
- Assembling synthetic biology circuits, metabolic pathways, and standardized DNA parts
Limitations and Practical Challenges
Cloning is robust but not trouble-free. Genes whose products are toxic to the host can kill the very cells meant to amplify them, requiring tightly controlled expression vectors or specialized strains. Very large inserts and sequences rich in repeats or strong secondary structure are unstable and can be deleted or rearranged by the host. Fragments amplified by PCR can acquire point mutations unless a high-fidelity polymerase is used, so sequence verification of the final construct is essential rather than optional.
Method-specific limitations also matter. Classical restriction–ligation leaves scar sequences and fails when the only available restriction sites also occur inside the insert. Seamless and recombination-based methods reduce these problems but add design overhead and reagent cost, and high-fragment assemblies still have a failure rate that grows with complexity. These are practical engineering constraints rather than fundamental barriers, and improved enzymes, standardized parts, and laboratory automation continue to narrow them.
Frequently Asked Questions
What is the difference between molecular cloning and PCR? PCR amplifies a DNA sequence by repeated enzymatic copying in a test tube, producing many linear copies but no living host or stable construct. Molecular cloning inserts a DNA fragment into a self-replicating vector and propagates it inside a host cell, yielding a stable, sequence-verified construct that can be stored, expressed, and reused. PCR is often used as an early step within a cloning workflow to generate the fragment to be cloned.
Is molecular cloning the same as cloning an animal? No. Molecular cloning copies a specific DNA fragment, such as a single gene, inside microbial or cultured cells. Reproductive cloning, as with Dolly the sheep, creates a whole organism that is genetically identical to a donor. The two share the word “clone” because both produce identical copies, but they operate at completely different scales and use different techniques.
Why is E. coli used in molecular cloning? Escherichia coli grows quickly, is inexpensive to culture, takes up foreign plasmids readily, and has been engineered into laboratory strains optimized for stable DNA maintenance. A single overnight culture started from one transformed cell can yield billions of identical plasmid copies, which makes E. coli the default host for amplifying and storing cloned DNA before it is moved to other systems.
What is the difference between a cloning vector and an expression vector? A cloning vector is designed mainly to carry, amplify, and stably maintain a DNA insert in a host cell. An expression vector additionally contains regulatory elements such as promoters, ribosome-binding sites in bacteria, Kozak sequences in eukaryotes, terminators, or polyadenylation signals that drive production of RNA or protein from the inserted gene. Many workflows clone a fragment into a simple cloning vector first and then transfer it into an expression vector.
Do you need restriction enzymes for molecular cloning? Not always. Classical cloning relies on restriction enzymes to cut DNA at defined sites, while many modern methods use overlap-based assembly, recombination, or type IIS restriction enzymes to reduce traditional restriction-site constraints. Restriction-enzyme cloning is still widely taught and used for simple, single-fragment constructs, and type IIS restriction enzymes remain central to Golden Gate cloning.
Is molecular cloning the same as gene editing? No. Molecular cloning builds and copies DNA constructs, often in bacteria or other host cells. Gene editing changes DNA sequences inside living cells or organisms. In practice, cloning is often used to build the plasmids or viral vectors that deliver gene-editing tools.
Further Reading
Proceedings of the National Academy of Sciences, Biochemical Method for Inserting New Genetic Information into DNA of Simian Virus 40
Proceedings of the National Academy of Sciences, Construction of Biologically Functional Bacterial Plasmids In Vitro
Nature Reviews Molecular Cell Biology, Bricks and Blueprints: Methods and Standards for DNA Assembly
