| Jul 07, 2026 |
Self-propelled nanoparticles hunt down target proteins inside tumor cellsSelf-propelled nanoparticles use tumor cell chemistry to find target proteins more efficiently and trigger their removal. |
| (Nanowerk Spotlight) Many medicines work by blocking a protein’s activity. They bind to a specific site, interfere with a signal, or slow an enzyme down. That strategy can be powerful, but it leaves the protein in place. In cancer and other diseases, some proteins drive harmful behavior not through a single vulnerable site, but through scaffolding roles, binding interactions, or persistent signaling states. In those cases, the more effective strategy may be to remove the protein altogether. |
| Removing a protein calls for a different kind of drug action. Targeted protein degraders do not try to keep a protein quiet. They recruit the cell’s disposal system. One major class, called PROTACs, short for proteolysis-targeting chimeras, works by chemically forcing a temporary meeting between the unwanted protein and an E3 ligase, an enzyme that helps attach ubiquitin tags. Those tags send the protein to the proteasome, where the cell breaks it down. |
| The chemistry only works when the right molecules meet often enough. A degrader has to find the target protein, bring it together with the tagging enzyme, and repeat that sequence inside a crowded cell. Nanoparticle versions can help degraders survive in the body, accumulate in tumors, and enter cells. Related work on nanoparticle-mediated targeted protein degradation has shown why nanoscale carriers are becoming important in this field. After entry, however, a passive particle still drifts until chance brings it near enough target proteins. |
| A study in Advanced Materials ("Autonomously Motile Nano‐PROTACs Act as Protein‐Sweeping Robots to Enhance Targeted Protein Degradation") addresses that hidden search problem by building motion into nanoparticle PROTACs. The researchers made gold nano-PROTACs, nanoscale versions of these degraders, with two different faces. One side carried ligands that bind the target protein and recruit the E3 ligase. The other carried catalase, an enzyme that converts hydrogen peroxide into oxygen and water. In the tumor cells tested, elevated hydrogen peroxide supplied the fuel for motion. |
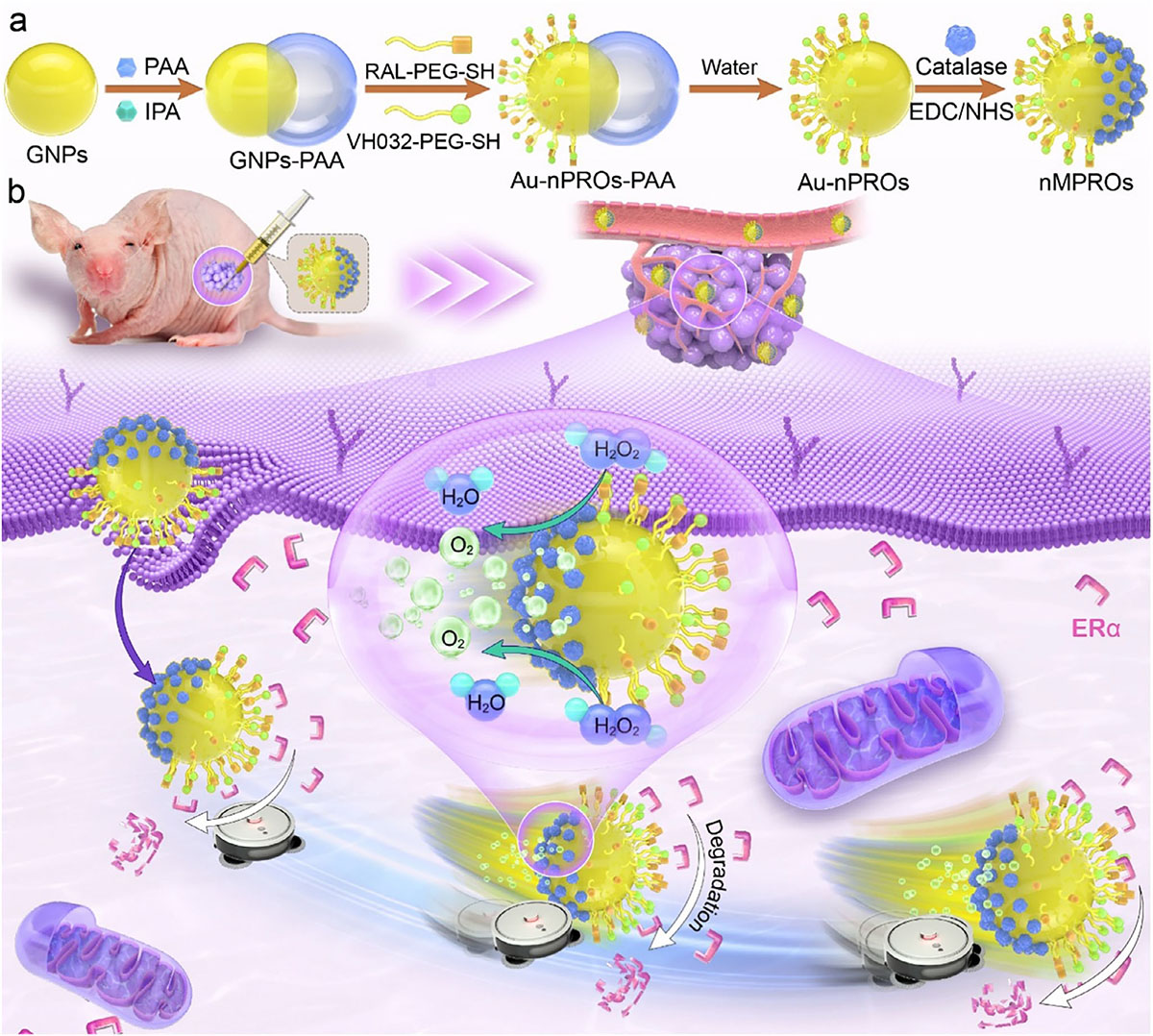 |
| Schematic diagram of nano-motoring PROTACs formulations and underlying therapeutic mechanisms. (Image: Reproduced with permission from Wiley-VCH Verlag) (click on image to enlarge) |
| The moving particles were no longer just carriers for degradation chemistry. They degraded estrogen receptor α with about threefold higher potency than static particles. By changing the target-binding ligand, the same design also redirected degradation toward PD-L1, an immune checkpoint protein studied in cancer therapy. Movement became part of how the system found and removed its target. |
| Hydrogen peroxide could drive motion only if the particle surface broke symmetry. Catalase spread evenly around a nanoparticle would still break down hydrogen peroxide, but the reaction would not favor one direction. Separating the functions across the gold surface solved that problem. The degradation ligands occupied one region, while catalase occupied the opposite region, allowing oxygen generation to bias movement. |
| Similar active-particle concepts appear in hydrogen peroxide-powered nanomotors in biological environments, though here the motion serves protein removal rather than clot treatment. |
| The same encounter problem appeared at the molecular scale. Raloxifene bound estrogen receptor α, while VH032 recruited the VHL E3 ligase. Flexible PEG chains tethered both molecules to the gold surface. Shorter, more rigid linkers might seem useful because they pull components closer together, but they weakened degradation. The recruited proteins needed room to adjust before ubiquitin tagging could occur. |
| That linker result sharpened the logic of the whole design. Productive degradation is not just about placing the right molecules nearby. The target protein and E3 ligase must also meet in a workable orientation. Flexible surface ligands helped form that local arrangement. Whole-particle propulsion addressed the larger search problem inside the cell, where passive diffusion limits how often useful encounters occur. |
| In breast cancer cells used as the main estrogen receptor model, static particles reproduced the core PROTAC mechanism. Estrogen receptor α fell only when the particles carried both the target-binding ligand and the E3-recruiting ligand. MG132, which blocks the proteasome, prevented that loss. Ubiquitin tags accumulated on estrogen receptor α after treatment. The particles were not simply stressing cells. They were routing the target into the intended disposal pathway. |
| Higher doses exposed the weakness of a passive, multivalent platform. At elevated concentrations, degradation declined instead of continuing to improve, a behavior the paper attributes to the hook effect. Too many binding sites can separate the target protein and E3 ligase into unproductive assemblies. The right pieces remain present, but the useful three-part complex becomes less frequent. |
| Catalase-bearing particles behaved differently when hydrogen peroxide was available. In solution, they consumed the fuel, generated oxygen, and moved farther as the concentration increased. Without hydrogen peroxide, they mostly drifted by Brownian motion. Free catalase mixed with static nano-PROTACs did not reproduce the behavior. Propulsion required catalase to be fixed to the particle in an asymmetric layout, not merely present nearby. |
| The cancer cells supplied the same fuel. The study compared breast cancer and melanoma cell models with normal endothelial cells, which line blood vessels. The cancer cells contained more hydrogen peroxide. After motile nano-PROTACs entered the breast cancer cells, intracellular hydrogen peroxide levels fell, consistent with catalase activity inside the cell. The chemistry that moved the particles in solution could therefore operate where degradation needed more efficient searching. |
| Comparable uptake made the performance difference more meaningful. Measurements of intracellular gold showed similar internalization among the relevant formulations. The motile particles nevertheless lowered estrogen receptor α more efficiently. Their half-maximal degradation concentration dropped to 2.153 µg/mL, compared with 6.649 µg/mL for the non-motile version. More particles were not entering the cells. The particles already inside were producing more degradation. |
| When hydrogen peroxide was depleted, much of the advantage disappeared. Pretreating cells with a hydrogen peroxide scavenger narrowed the performance gap between motile and static nano-PROTACs. Without proteasome activity, estrogen receptor α returned. After treatment, ubiquitin tags increased on the target protein. The controls point to motility-enhanced proteasomal degradation rather than nonspecific protein loss. |
| PD-L1 gave the design a different test. The researchers replaced raloxifene with BMS-8, which binds PD-L1, while keeping the catalase motor architecture. The motor remained active after the ligand swap. In melanoma cells, the motile nano-PROTACs degraded about 85 % of PD-L1, outperforming static particles and catalase mixed with static particles. Motion still helped after the target biology changed. |
| The first animal experiment stayed local. The researchers injected the particles directly into breast tumors in mice rather than through the bloodstream. Under those conditions, motile particles remained detectable in tumor tissue longer than free dye, with a tumor half-life of about 27 hours. Across the 14 day treatment period, they suppressed tumor growth more strongly than static nano-PROTACs. |
| That result gives the platform an early in vivo signal, not a finished therapeutic case. Treated mice showed no major body-weight loss, and blood analysis plus tissue staining of major organs did not reveal obvious acute damage under the tested conditions. Local injection leaves harder questions open, including bloodstream delivery, biodistribution, clearance, immune response, repeated dosing, and scalable manufacturing. |
| The important advance lies inside the cell. Nanoparticles are often judged by whether they can reach a tissue and enter a cell. For targeted protein degradation, entry is only the start. A passive nano-PROTAC waits for the right encounter in a crowded environment. A motile one can sample more of that environment and improve the chance that the target protein, the E3 ligase, and the degrader meet correctly. |
| Future protein-removing nanomedicines may need to control delivery, local binding geometry, and intracellular movement together. This paper shows why those layers cannot be separated easily. Flexible ligands help assemble the degradation complex once contact occurs. Autonomous motion helps create more chances for that contact. Inside the cell, finding the protein can be as important as binding it. |
 By
Michael
Berger
– Michael is author of four books by the Royal Society of Chemistry:
Nano-Society: Pushing the Boundaries of Technology (2009),
Nanotechnology: The Future is Tiny (2016),
Nanoengineering: The Skills and Tools Making Technology Invisible (2019), and
Waste not! How Nanotechnologies Can Increase Efficiencies Throughout Society (2025)
Copyright ©
Nanowerk LLC
By
Michael
Berger
– Michael is author of four books by the Royal Society of Chemistry:
Nano-Society: Pushing the Boundaries of Technology (2009),
Nanotechnology: The Future is Tiny (2016),
Nanoengineering: The Skills and Tools Making Technology Invisible (2019), and
Waste not! How Nanotechnologies Can Increase Efficiencies Throughout Society (2025)
Copyright ©
Nanowerk LLC
|
|
ORCID information
|

Become a Spotlight guest author! Join our large and growing group of guest contributors. Have you just published a scientific paper or have other exciting developments to share with the nanotechnology community? Here is how to publish on nanowerk.com. |
